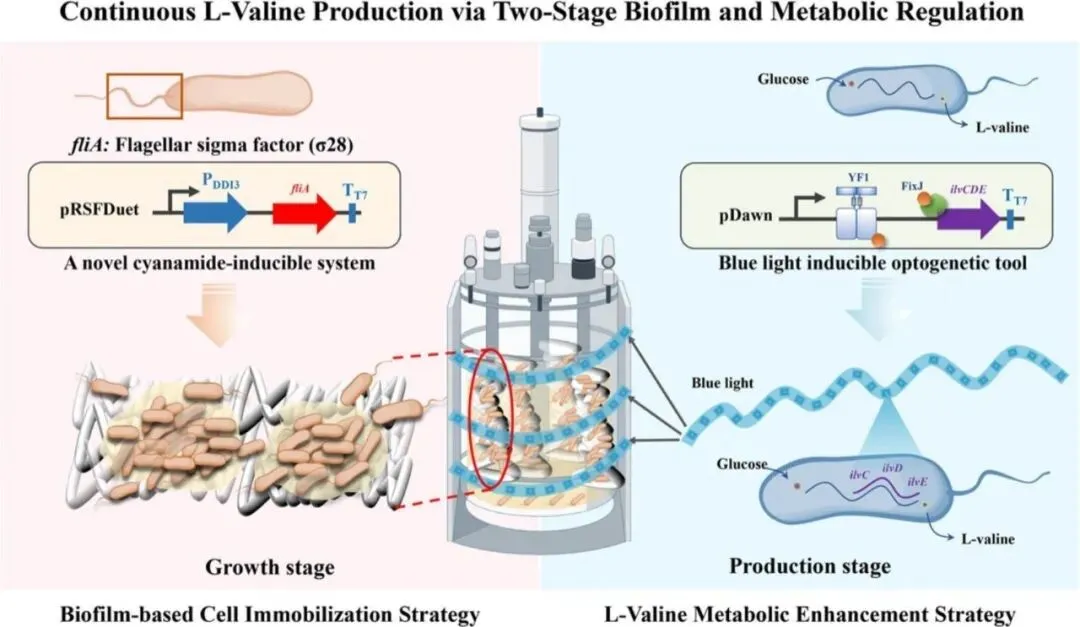
L-缬氨酸作为支链氨基酸家族的重要成员,在饲料、医药和食品工业中具有广泛应用。然而,由于代谢途径调控欠佳和细胞 retention 不足,实现氨基酸的连续微生物发酵仍面临巨大挑战。近日,南京工业大学应汉杰院士团队与孙文俊副研究员在《Chemical Engineering Journal》发表研究,通过构建增强型生物被膜形成与蓝光激活代谢的工程化大肠杆菌,成功实现了L-缬氨酸的高产长效发酵。研究首先通过探究鞭毛及运动性相关基因对生物被膜形成的影响,构建了新型氰胺诱导系统(DDI3启动子)在生长阶段诱导fliA表达,加速生物被膜形成;随后在生产阶段通过光遗传学工具pDawn瞬时激活ilvCDE基因表达,显著提升L-缬氨酸代谢通量。最终工程菌株PDDI3-fliA+pDawn-ilvCDE在50 L生物反应器中实现462小时连续发酵,L-缬氨酸产量达71.63 g/L,葡萄糖转化率达0.57 g/g,生产强度达2.05 g/L/h。据悉,这是迄今报道的大肠杆菌中葡萄糖转化L-缬氨酸的最高产率。该研究为L-缬氨酸的连续发酵提供了系统性的代谢工程策略,并可推广至其他代谢产物的生物合成摘要
由于代谢途径控制欠佳和细胞保留不足,氨基酸的连续微生物发酵一直面临着巨大的挑战。L-缬氨酸是一种必需氨基酸,对饲料和制药等多个行业至关重要,但其在连续条件下的高效、持续生物合成却鲜有实现。
本研究构建了一种具有增强生物膜形成能力和蓝光激活代谢的工程化大肠杆菌(E. coli),用于高产、长期发酵。通过研究鞭毛和运动相关基因,我们开发了一种新型的氰胺诱导系统,该系统携带DDI3启动子,可诱导fliA基因表达,从而加速生长阶段的生物膜形成。随后,为了提高L-缬氨酸的代谢通量,我们评估了ilvCDE的表达,并在生产阶段利用光遗传工具pDawn进行瞬时激活,显著提高了其催化效率。采用两阶段调控策略,最终菌株 P DDI3 -fliA+pDawn-ilvCDE 在 50 L 生物反应器中连续发酵 462 小时,L-缬氨酸产量达到 71.63 g/L,葡萄糖转化为 L-缬氨酸的转化率为 0.57 g/g,生产率为 2.05 g/L/h。据我们所知,这是迄今为止报道的大肠杆菌中最高的葡萄糖转化为 L-缬氨酸的转化率。这种系统的代谢工程策略为实现 L-缬氨酸的连续发酵建立了一个前景广阔且用途广泛的平台,并可轻松扩展到其他代谢物的生物合成。
 研究背景与意义
研究背景与意义
L-缬氨酸作为人体必需支链氨基酸,在饲料添加、医药合成及功能食品等领域具有重要应用价值。凭借其可持续性和环境友好优势,微生物发酵已成为L-缬氨酸生产的主流方法。
高效微生物发酵常依赖于细胞的大规模聚集,进而形成生物被膜。特别是具有高耐受性的生物被膜相比浮游细胞更能抵御恶劣环境。已有研究表明,通过过表达fimH基因可增强大肠杆菌W1688的生物被膜形成量,使L-苏氨酸产量在分批发酵中从10.5 g/L提高至14.1 g/L,并在连续发酵中进一步增至17.5 g/L。同样,过表达PotD蛋白也能刺激大肠杆菌BL21(DE3)的生物被膜形成。
稳健的生物被膜形成保证了强大且高产细胞工厂的建立,进而提升整体过程的生产力。与此同时,代谢工程策略为L-缬氨酸的高效外排提供了广阔前景。虽然这些研究取得了可观进展,但多数策略受限于分批处理,尚未能提供长期稳定生产L-缬氨酸的可持续解决方案,并且细胞生长与L-缬氨酸生物合成之间存在权衡关系,当此平衡被打破时,丙酮酸分配将偏向核心代谢途径,过度强调产物合成可能导致生物量不足,降低生产强度和产率。
本研究针对上述挑战,开发了一种两阶段工程策略,将生物被膜介导的固定化与靶向代谢途径强化相结合,为实现L-缬氨酸连续发酵提供了创新解决方案。
 研究方法与技术路线
研究方法与技术路线2.1 菌株、质粒与培养条件
本研究所用菌株和质粒汇总于表S1和S2,引物序列见表S3。采用pCas和pTarget质粒进行基因组编辑,pTrc99a、pRSFDuet、pACYCDuet和pDawn用于基因表达。大肠杆菌045菌株作为L-缬氨酸生产菌株和遗传改造的出发菌株。大肠杆菌在LB培养基中37°C培养,添加异丙基-β-D-硫代半乳糖苷(IPTG)、氰胺和蓝光诱导基因表达。
2.2 工程菌株构建
采用CRISPR-Cas9辅助同源重组进行基因敲除。针对目标基因设计sgRNA克隆至pTarget,共转化携带同源臂的供体DNA片段进行编辑。阳性克隆经菌落PCR筛选并经Sanger测序确认。根据标准分子克隆技术,将相应编码序列插入pTrc99a和pRSFDuet质粒构建氰胺诱导系统。同样,采用pACYCDuet和pDawn质粒用于光遗传学介导的L-缬氨酸代谢系统调控。为评估鞭毛运动性对生物被膜形成的影响,构建过表达motA+B、flgM、fliA、flgK+L等的菌株。合成启动子PDDI2和PDDI3用于pRSFDuet衍生载体中的可调表达,光诱导表达通过pDawn系统实现。
2.3 生物量、葡萄糖浓度和L-缬氨酸浓度测定
生物量以600 nm处吸光度(OD600)定量。葡萄糖浓度采用3,5-二硝基水杨酸法测定。L-缬氨酸浓度采用高效液相色谱分析,流动相为乙腈/水(50:50, v/v)和50 mM乙酸钠,流速0.6 mL/min,紫外检测波长360 nm。
2.4 生长曲线测定与生物被膜定量
大肠杆菌在不同氰胺浓度和光照条件下的生长曲线按前期方法测定。将过夜培养的大肠杆菌稀释至初始OD600 0.1,每2小时用紫外-可见分光光度计测定OD600。氰胺浓度分别设为10 mM、20 mM和30 mM。为研究蓝光对细胞生长的影响,培养物暴露于不同蓝光强度(300 lx、600 lx和900 lx)或完全避光作为对照。生物被膜定量采用结晶紫染色法。
2.5 运动性测定
采用游动和群集运动性测定评估软琼脂平板上的运动表型。群集运动性在群集琼脂平板(含10 g/L蛋白胨、5 g/L酵母提取物、10 g/L NaCl、3 g/L葡萄糖和3 g/L Eiken琼脂)上进行,将1.2 μL细菌培养物点样于平板中心,25°C静置培养16小时。游动运动性在游动琼脂平板(含10 g/L蛋白胨、7 g/L NaCl和5 g/L Eiken琼脂)上进行,用无菌接种针穿刺接种,25°C静置直立培养40小时。
2.6 工程菌鞭毛与生物被膜的可视化分析
采用扫描电子显微镜观察大肠杆菌菌株在玻璃片和棉纤维上的表面附着差异。SEM也用于评估大肠杆菌细胞的形态完整性。经适当固定和脱水处理,细菌悬浮液进行细胞切片处理。采用透射电子显微镜采集鞭毛图像,加速电压80-120 kV。为可视化工程菌的生物被膜形成,采用4',6-二脒基-2-苯基吲哚(DAPI)染色细胞,用蔡司LSM880共聚焦激光扫描显微镜成像。
2.7 相对荧光强度分析与实时定量PCR
对4株大肠杆菌进行相对荧光强度分析:Ptrc-eGFP、PDDI2-eGFP、PDDI3-eGFP和野生型大肠杆菌045。各菌株在LB培养基中37°C、220 rpm过夜培养。其中,Ptrc-eGFP菌株用0.5 mM IPTG诱导,PDDI2-eGFP和PDDI3-eGFP菌株用不同浓度氰胺(10 mM、20 mM、30 mM)诱导。将细胞悬液OD600稀释至0.5后,在激发波长490 nm、发射波长530 nm处测定荧光强度。相对荧光强度按公式计算。
采用实时定量PCR定量鞭毛运动相关基因(motA、motB、flgM、flgN、fliA、flgK、flgL)和L-缬氨酸生物合成途径基因(ilvC、ilvD、ilvE、ilvI、ilvH)的转录水平。使用Vazyme细菌RNA提取试剂盒提取总RNA,用HiScript® II Q RT SuperMix合成cDNA,qRT-PCR使用AceQ® qPCR SYBR® Green Master Mix进行,以16S rRNA为内参,采用2-ΔΔCT法计算相对表达量。
2.8 两阶段调控工程菌的连续发酵
种子培养和游离细胞发酵的培养基组成参考文献。将2 L种子培养液转接至50 L锯齿状生物反应器。初始发酵在37°C进行,通气量1.3 m³/h,搅拌转速600 rpm,罐压0.05 MPa。通过补加氨水维持pH约7.0。每2小时取发酵液监测葡萄糖浓度和OD600。当残糖降至约1 g/L时,补加600 g/L葡萄糖溶液维持葡萄糖浓度在1-2 g/L。当OD600达到40-50时,发酵转入催化阶段,搅拌和通气减半,同时确保溶氧维持在25%以上。当可见L-缬氨酸晶体(80-90 g/L)开始形成时终止发酵。
对于固定化连续发酵,选择棉纤维作为固定化载体,以之字形固定在反应器内的载体框架上。发酵和补料条件与分批过程一致。每批结束后移除发酵液,保留带有生物被膜的棉纤维,随后泵入新鲜无菌发酵培养基进行下一周期,实现L-缬氨酸的连续发酵。
对于蓝光发酵系统,商用蓝色LED灯带(峰值波长460 nm)环绕在生物反应器外壁。为提高照射均匀性,棉纤维垂直安装并与反应器壁平行排列,使LED光源到生物被膜表面的光程距离固定为2 cm。光照强度通过量子计测定,范围为50-128 μmol/m²/s,平均75.5 μmol/m²/s。
2.9 统计分析
所有实验至少重复三次。采用GraphPad Prism软件进行单因素或双因素方差分析或配对t检验评估统计学显著性。数据以三次独立实验的均值和标准差表示。
 结果与讨论
结果与讨论
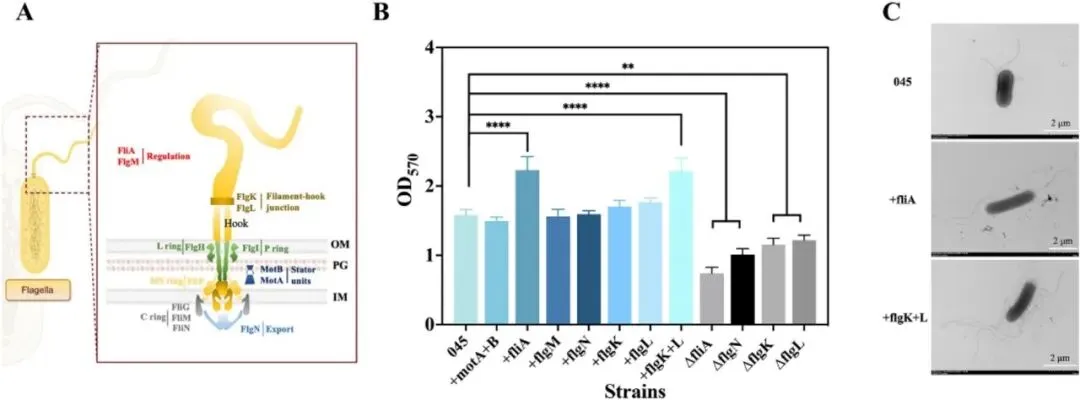
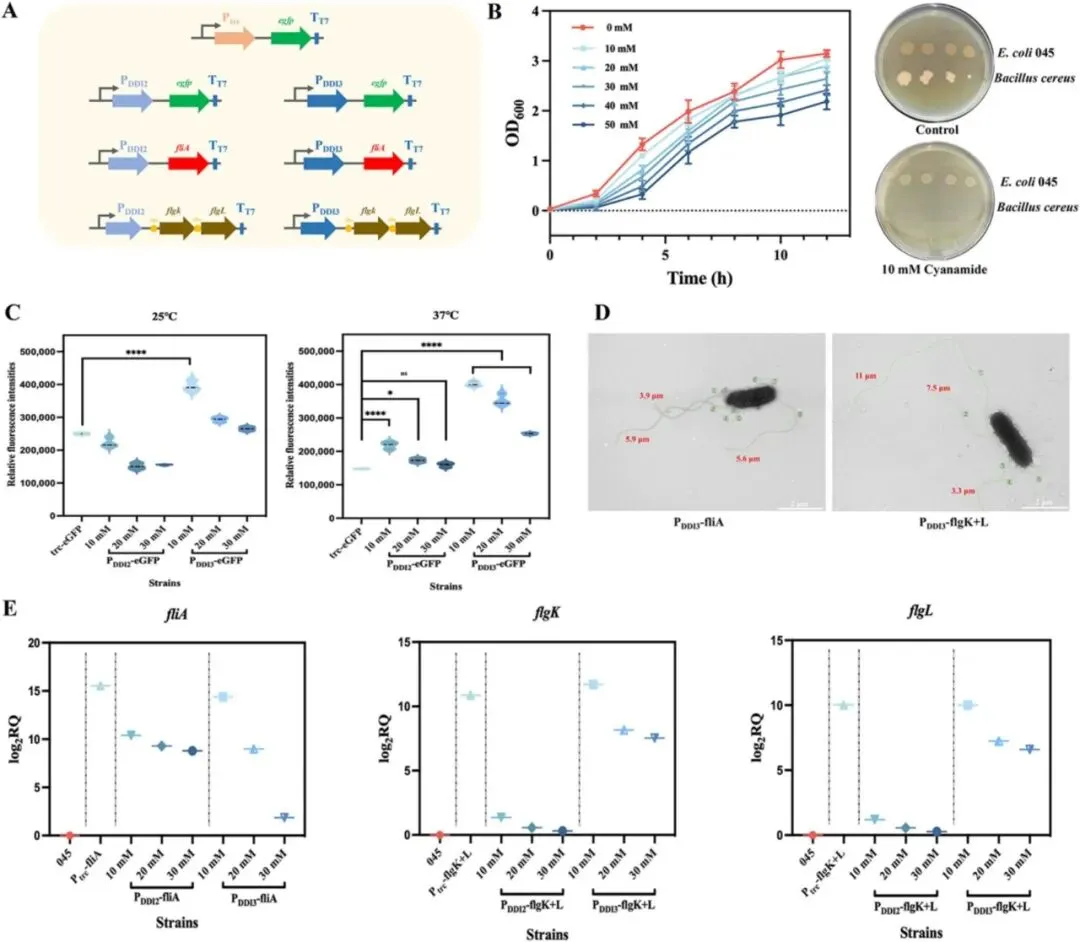
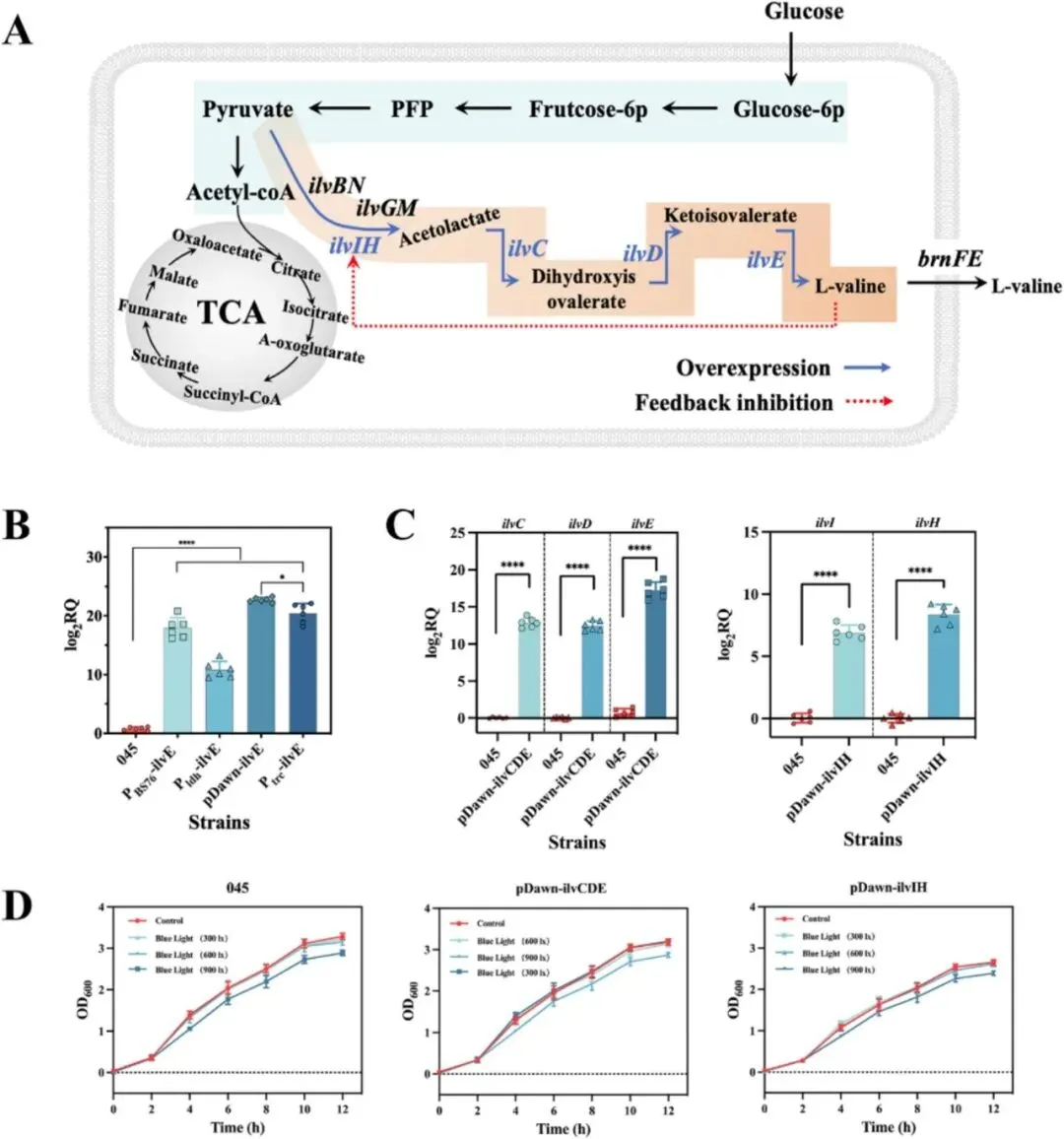
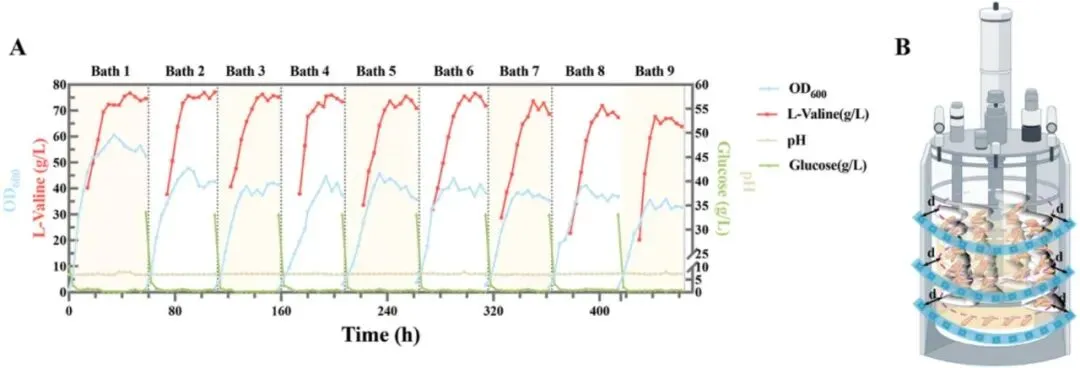
3.1 鞭毛运动性基因对大肠杆菌生物被膜形成的影响如图1A所示,研究选取一组鞭毛及运动性相关基因,包括motA、motB、fliA、flgM、flgN、flgK和flgL进行遗传操作,探究其在大肠杆菌生物被膜形成中的作用。为增强生长早期至生物被膜发育阶段的细菌运动性,通过质粒表达构建了过表达菌株motA+B、flgM、fliA、flgN、flgK、flgL和flgK+L。同时采用CRISPR-Cas9系统构建了fliA、flgN、flgK和flgL的敲除菌株,以阐明鞭毛功能和运动性对生物被膜形成的影响。结果显示,与出发菌株大肠杆菌045相比,+fliA和+flgK+L菌株的生物被膜形成显著增强,分别提高了41.14%和39.87%,而敲除菌株呈现相反趋势(图1B)。图1 鞭毛相关基因对大肠杆菌运动性和生物被膜形成的调控通过TEM分析各菌株的鞭毛形态差异和数量(图1C)。与大肠杆菌045相比,鞭毛数量明显增加。+flgK+L菌株不仅表现出更多鞭毛,鞭毛长度也显著增加,证实fliA和flgK+L在鞭毛表型中发挥重要调控作用。游动和群集运动性测定表明,与大肠杆菌045不同,motA+B、fliA和flgK+L的过表达显著增强了细胞运动性,而其他基因影响甚微。运动性对大肠杆菌生物被膜形成的起始至关重要。大肠杆菌的鞭毛为细胞提供向表面运动所需的运动性,驱动初始粘附,从而促进生物被膜形成。鉴于鞭毛运动相关基因调控对生物被膜形成的积极影响,研究构建了新型氰胺诱导表达系统以进一步增强生物被膜发育(图2A),同时可在生长阶段降低杂菌污染风险。尽管氰胺具有一定细胞毒性,但OD600的波动表明10 mM氰胺对细胞生长影响最小(图2B)。随着氰胺浓度升高,其对大肠杆菌生长增殖的抑制作用更为明显。值得注意的是,10 mM氰胺诱导对主要污染源蜡样芽孢杆菌的抑制作用显著且明显。为成功构建氰胺诱导表达系统,将酿酒酵母来源的DDI2和DDI3启动子经密码子优化后在大肠杆菌中表达。令人惊讶的是,两个DDI启动子均表现出温度非依赖性诱导,这对大肠杆菌所需的恒温发酵有利。相比之下,以Ptrc-eGFP菌株为对照,25°C和37°C下的荧光强度结果表明trc启动子呈温度依赖性(图2C)。此外,DDI3启动子在10 mM氰胺下诱导效率最高,与图2B趋势一致。通过定量fliA、flgK和flgL的表达进一步证实,DDI3启动子在选定诱导条件下有效驱动这些基因的转录(图2E)。值得注意的是,DDI3启动子的引入不仅增加了鞭毛数量,还延长了鞭毛长度,PDDI3-fliA和PDDI3-flgK+L菌株的生物被膜形成分别提高了54.7%和68.8%(图2D)。SEM和CLSM图像显示,通过过表达fliA和flgK+L介导的鞭毛运动性增强了细胞聚集,形成更致密、更有结构的生物被膜。总体而言,稳健的细胞聚集和生物被膜形成与强化发酵和产物转化密切相关。L-缬氨酸产量测定显示,大肠杆菌045、PDDI3-fliA和PDDI3-flgK+L分别为84.42 g/L、87.35 g/L和65.64 g/L。PDDI3-fliA的生产强度(2.56 g/L/h)和葡萄糖转化率(0.46 g/g)也略高于大肠杆菌045(2.23 g/L/h和0.42 g/g)和PDDI3-flgK+L(1.97 g/L/h和0.32 g/g)。值得注意的是,工程菌PDDI3-fliA比大肠杆菌045更早转入生产阶段,这可能有助于缩短发酵周期并提高生产力。相反,PDDI3-flgK+L的L-缬氨酸产量大幅下降,可能归因于工程改造导致的代谢平衡显著破坏。3.3 蓝光激活L-缬氨酸代谢的光遗传学调控及两阶段策略在连续发酵中的应用除在生长阶段构建氰胺诱导表达系统促进生物被膜形成外,研究还通过蓝光激活进一步调控L-缬氨酸代谢通量。根据L-缬氨酸的主要生物合成途径(图3A),葡萄糖作为唯一碳源,被大肠杆菌吸收后通过糖酵解途径代谢产生丙酮酸,经系列酶促反应最终转化为L-缬氨酸。其中关键酶包括乙酰羟酸合成酶III(AHAS III,由ilvIH编码)、乙酰羟酸异构还原酶(ilvC)、二羟酸脱水酶(ilvD)和转氨酶(ilvE)。为确保高效酶表达,以ilvE为报告基因筛选组成型启动子PBS76、厌氧启动子Pldh和蓝光诱导光遗传学质粒pDawn(图3B)。尽管原有的两阶段发酵过程涉及有氧生长阶段后厌氧催化,但研究发现蓝光诱导的pDawn质粒使ilvE表达量最高。值得注意的是,尽管trc启动子具有高诱导表达特性,但ilvE的表达量(0.5 mM IPTG)明显低于pDawn。在蓝光诱导下,L-缬氨酸代谢途径关键基因(ilvC、ilvD、ilvE、ilvH、ilvI)的表达均显著增强(图3C),有力验证了光遗传学调控在L-缬氨酸代谢途径中的可行性。进一步评估不同光强下pDawn-ilvCDE和pDawn-ilvIH菌株的生长情况(图3D),在光强低于900 lx时,pDawn-ilvCDE与大肠杆菌045生长趋势相似,而pDawn-ilvIH出现生长抑制,可能归因于ilvIH过表达导致的代谢失衡或辅因子耗竭。在各测试基因中,ilvCDE被证实效果最佳,并被选用于增强L-缬氨酸生产。这一结论也通过代谢工程菌株pDawn-ilvCDE和pDawn-ilvIH的游离细胞发酵实验得到验证。在发酵36小时,大肠杆菌045、pDawn-ilvCDE和pDawn-ilvIH的L-缬氨酸产量分别达到85.12 g/L、90.87 g/L和48.52 g/L。其中,pDawn-ilvCDE菌株的平均生产强度和葡萄糖转化率分别为2.41 g/L/h和0.44 g/g,高于大肠杆菌045和pDawn-ilvIH。出乎意料的是,pDawn-ilvIH表现出L-缬氨酸产量和细胞生长的显著下降。在600 lx光强下进一步明确诱导时间,提前2小时诱导(从14小时调至12小时)使L-缬氨酸产量提高13.92%。基于生长阶段氰胺诱导表达系统和生产阶段代谢途径精细调控的构建,最终将两阶段调控策略整合应用于L-缬氨酸连续发酵。具体而言,构建了双质粒共表达菌株PDDI3-fliA+pDawn-ilvCDE(图4A),发酵生物反应器的物理设置如图4B所示。在整个连续发酵过程中,固定化的PDDI3-fliA+pDawn-ilvCDE保持高细胞活力和持续的L-缬氨酸生产能力,9个批次平均终产量达71.45 g/L(图4C)。与大肠杆菌045相比,PDDI3-fliA+pDawn-ilvCDE的L-缬氨酸产量提高13.00%,有助于缩短发酵周期并提高生产力。葡萄糖转化率和生产强度分别达到0.57 g/g和2.05 g/L/h。此外,PDDI3-fliA+pDawn-ilvCDE在发酵早期表现出更快的生长速率,在生产阶段更快达到70 g/L的L-缬氨酸浓度。同时,在连续9个批次共462小时的发酵过程中,过程保持稳定,证明了PDDI3-fliA+pDawn-ilvCDE的稳健性及其持续高产L-缬氨酸的潜力。表1全面比较了大肠杆菌045与两阶段调控策略工程菌株在L-缬氨酸生产方面的差异。尽管微生物发酵是L-缬氨酸生产的常用方法,但细胞生理状态的不稳定和代谢途径的相互依赖常阻碍L-缬氨酸的有效生产。借助鞭毛调控对大肠杆菌045生物被膜形成的积极作用,首先构建了新型氰胺诱导系统增强生物被膜发育。比较游离细胞发酵和固定化发酵模式,PDDI3-fliA菌株较大肠杆菌045实现了更高的L-缬氨酸产量(87.35 g/L)、产率(0.46 g/g葡萄糖)和生产强度(2.56 g/L/h)。随后,蓝光激活的引入进一步提升了工程菌株的性能,如pDawn-ilvCDE表现出90.87 g/L的产量、0.44 g/g葡萄糖的产率和2.41 g/L/h的生产强度,显著高于标准游离细胞发酵和蓝光调控的pDawn-ilvIH(48.08 g/L、0.26 g/g葡萄糖和1.56 g/L/h)。表1 本研究及文献中不同调控策略下起始菌株与工程菌株的L-缬氨酸生产比较目前报道的使用重组谷氨酸棒杆菌生产的最高L-缬氨酸浓度为86.5 g/L,产率0.234 g/g,而两株重组大肠杆菌VHY18和VAL38分别产L-缬氨酸84 g/L和92 g/L。与前述研究不同,为避免L-缬氨酸结晶干扰,本研究将最终L-缬氨酸浓度控制在约80 g/L以下。特别地,通过双质粒共表达菌株PDDI3-fliA+pDawn-ilvCDE在固定化发酵中结合氰胺诱导表达系统和代谢途径精细调控,显示出卓越的葡萄糖转化率0.57 g/g,这是迄今报道的如此长连续发酵时间下的最高转化效率。此外,两阶段调控策略不仅提高了整体L-缬氨酸产量,还提供了对代谢通量的更强控制和跨发酵周期的菌株稳定性增强。值得注意的是,与传统的化学诱导方法相比,蓝光调控在经济性上具有显著优势。在50 L生物反应器条件下对IPTG化学诱导和蓝光调控的运行成本进行定量比较发现,在9批连续发酵过程中,IPTG诱导成本约为蓝光调控相关电费的4.3倍(表S7)。该蓝光调控生物被膜介导发酵系统的关键放大原理是保持光源到生物被膜的恒定光程距离,而非试图照亮整个发酵液体积。通过将固定化载体垂直安装并与反应器壁平行排列,光源到生物被膜的距离固定为2 cm,实现了对生物被膜中高密度催化群体的可重复光调控。这种几何结构通过模块化外部LED灯带安装和载体面板倍增提供了一条实用的放大途径,同时保持了生物被膜表面的局部光子通量。与传统的发酵方法相比,氰胺诱导表达与蓝光诱导光遗传学调控的整合提供了一种创新且灵活的调控框架,可适用于其他高价值生化品的生产,使其成为工业生物技术中的优越替代方案。