3.1. 基因组测序、从头组装与功能注释
基因组测序:结合Illumina、PacBio SMRT和Hi-C技术对二倍体希腊牛至进行测序,预估基因组大小为732.76 Mb,杂合度为1.58%,重复序列占71.23%。
组装与评估:最终组装出709.74 Mb的染色体级别基因组,锚定至15条假染色体,支架N50达46.36 Mb;组装质量评估显示高比对率(98.94%)、高覆盖度(99.81%)及核心基因高完整度(96.20%)。
注释与挖掘:成功预测了30,891个蛋白编码基因,其中98.23%获得功能注释;同时鉴定了大量非编码RNA及302个OvCYP基因,并筛选出17个在突变体中差异表达的潜在萜类合成关键基因。
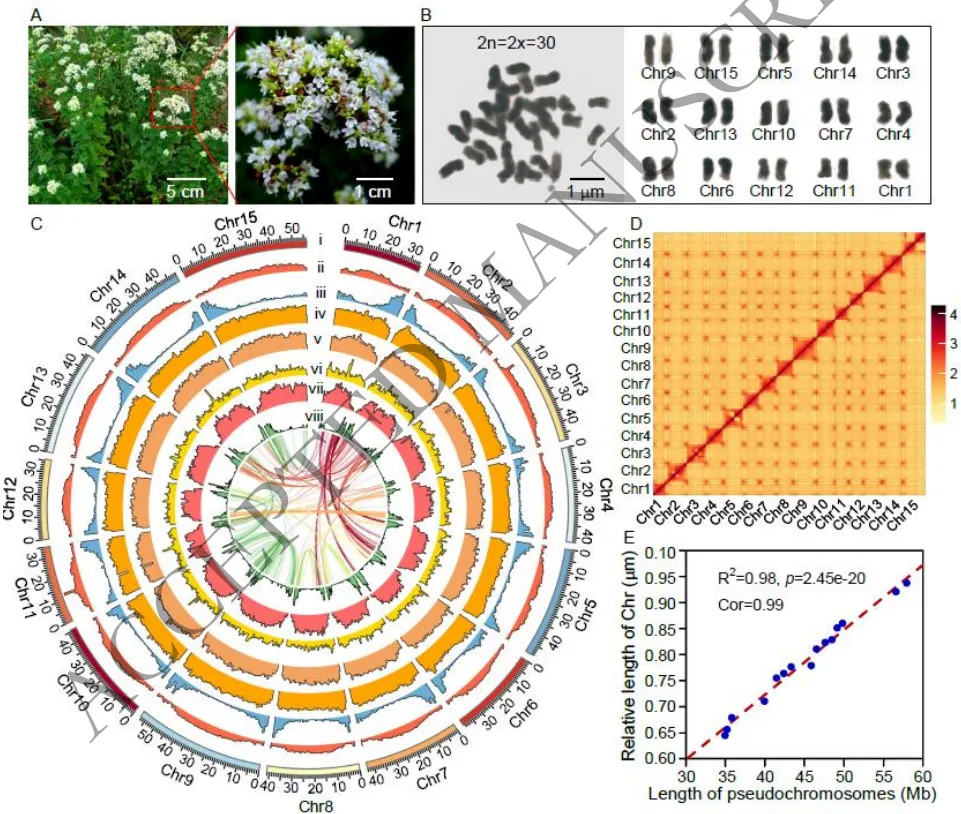
利用多平台测序技术成功构建了希腊牛至(2n=30)的高质量染色体级别基因组。组装结果显示基因组总长为709.74 Mb,其中94.77%的序列被锚定至15条假染色体上,支架N50达到46.36 Mb。该组装具有极高的准确性(Qv=40.46)和完整性(96.20%核心基因被找回)。基因组中包含70.84%的重复序列(主要为LTRs),并成功注释了30,891个蛋白编码基因。通过对细胞色素P450(CYP450)基因家族的深度挖掘,共鉴定出302个OvCYP基因,系统发育及表达模式分析进一步锁定了多个可能参与单萜和倍半萜生物合成的候选基因(Fig. 1)(Fig. S1-S7)。
图1. 牛至表型及染色体级别基因组组装特征。图A和B展示了二倍体希腊牛至植株及染色体计数;图C和D展示了15条假染色体的组装特征及长度分布;图E显示了组装长度与体细胞测量值的高度一致性。
3.2 比较基因组学分析
系统发育:基于14个植物物种的单拷贝直系同源基因构建进化树,明确了希腊牛至与同属唇形科的百里香亲缘关系最近,分化时间约为6.20百万年前。
基因扩增:比较基因组分析揭示了希腊牛至中存在314个显著扩增的基因家族,这些家族主要富集于次生代谢过程及特定酶活性调控等功能通路。
特异基因:鉴定出270个希腊牛至特异性基因家族,涉及837个基因,主要富集于FK506结合核蛋白及特定甲基转移酶等功能。
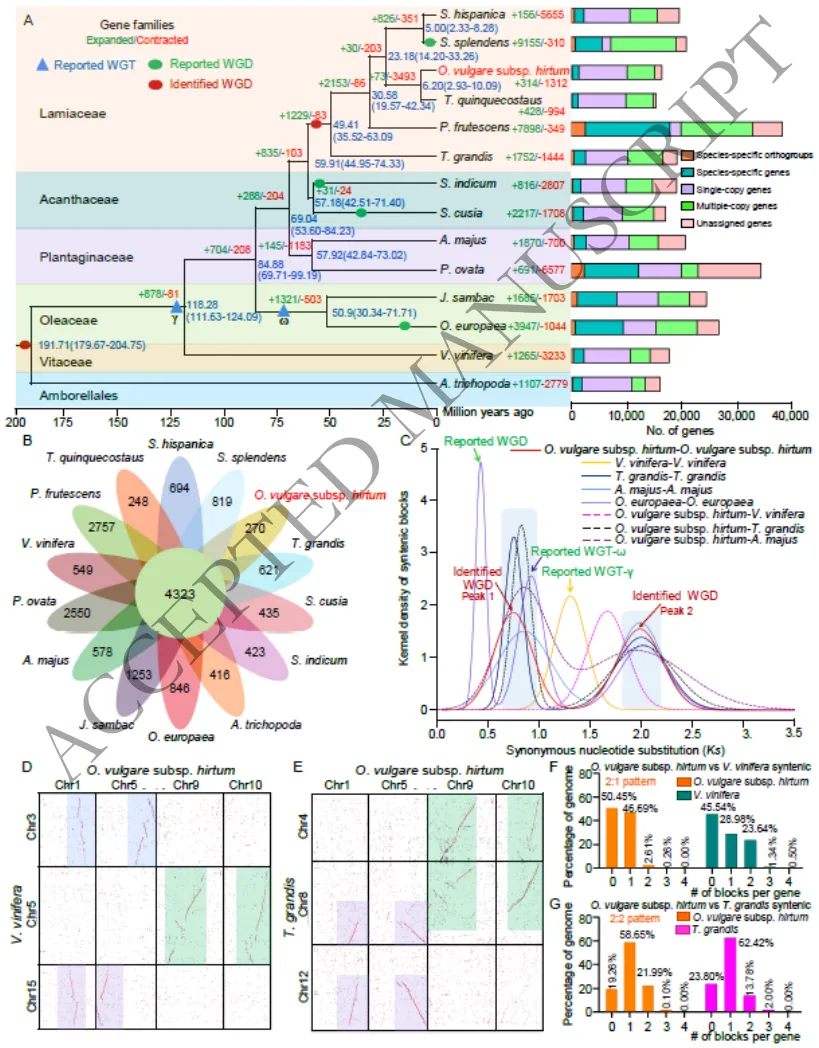
通过对包含葡萄(外群)在内的14个物种进行比较基因组学分析,精准重构了希腊牛至的进化地位。系统发育树表明,希腊牛至与百里香(Thymus quinquecostatus)的亲缘关系最为密切,二者约在6.20百万年前发生分化;而在更高分类层级上,唇形科与爵床科的亲缘关系最近。基因家族分析显示,希腊牛至基因组中存在314个显著扩增的基因家族,这些家族在次生代谢过程等功能中高度富集,为解析其独特的芳香特性提供了遗传线索。此外,还鉴定出270个物种特异性基因家族(Fig. 2A-C)(Fig. S8-S9)。
图2. 希腊牛至及代表性物种的进化、基因家族与全基因组复制(WGD)分析。图A展示了包含希腊牛至在内的14个代表性物种的系统发育关系及分化时间(如与百里香约在6.20百万年前分化);图B-C展示了扩增基因家族及物种特异性基因家族的分布情况,同时图C也显示了基于同义替换率(Ks)分布揭示的WGD事件发生时间;图D-G展示了希腊牛至与代表性物种(如柚木、葡萄等)之间的染色体共线性映射关系。
3.3 核型进化分析
全基因复制:揭示了希腊牛至经历了两次显著的全基因组复制(WGD)事件,其中较近的一次(约58.92百万年前)为与柚木共享的谱系特异性复制。
染色体共线:证实了希腊牛至与柚木、葡萄之间存在广泛的全基因组共线性,其中与葡萄呈现2:1的共线性比例,与柚木呈现2:2的共线性比例。
核型演化:重构了祖先唇形目核型(ALK),详细描绘了从21条祖先真双子叶植物原染色体经过融合、断裂及易位等事件,最终演化为希腊牛至15条染色体的复杂过程。
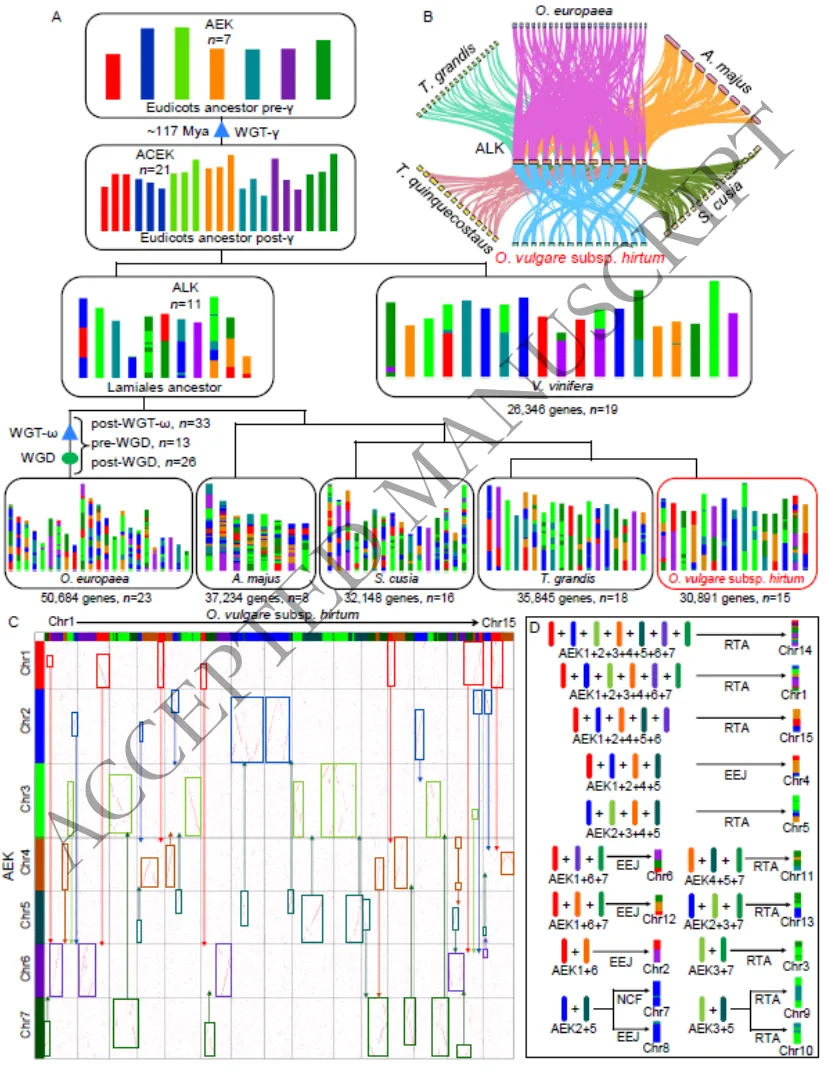
为探究希腊牛至的古进化历史,开展了深度的共线性与核型进化分析。同义替换率(Ks)分布揭示了希腊牛至基因组经历了两次全基因组复制(WGD)事件:一次是约185.31百万年前的古老复制,另一次是约58.92百万年前与柚木(Tectona grandis)共享的谱系特异性复制。广泛的基因组共线性分析证实了希腊牛至与葡萄之间存在2:1的同源映射关系。通过重构祖先唇形目核型(ALK),清晰地追溯了希腊牛至15条现代染色体的演化轨迹,表明其经历了复杂的嵌套染色体融合(NCF)、端对端连接(EEJ)及染色体臂相互易位(RTA)等重排事件(Fig. 2C-G)(Fig. 3)(Fig. S10-S11)。
图3.希腊牛至的核型进化轨迹。图A展示了从祖先唇形目核型(ALK)到现代物种的整体演化路径;图B-D详细描绘了从21条祖先原染色体演化为希腊牛至15条染色体过程中,所经历的染色体片段融合(NCF)、断裂、端对端连接(EEJ)及重排(RTA)等复杂事件。
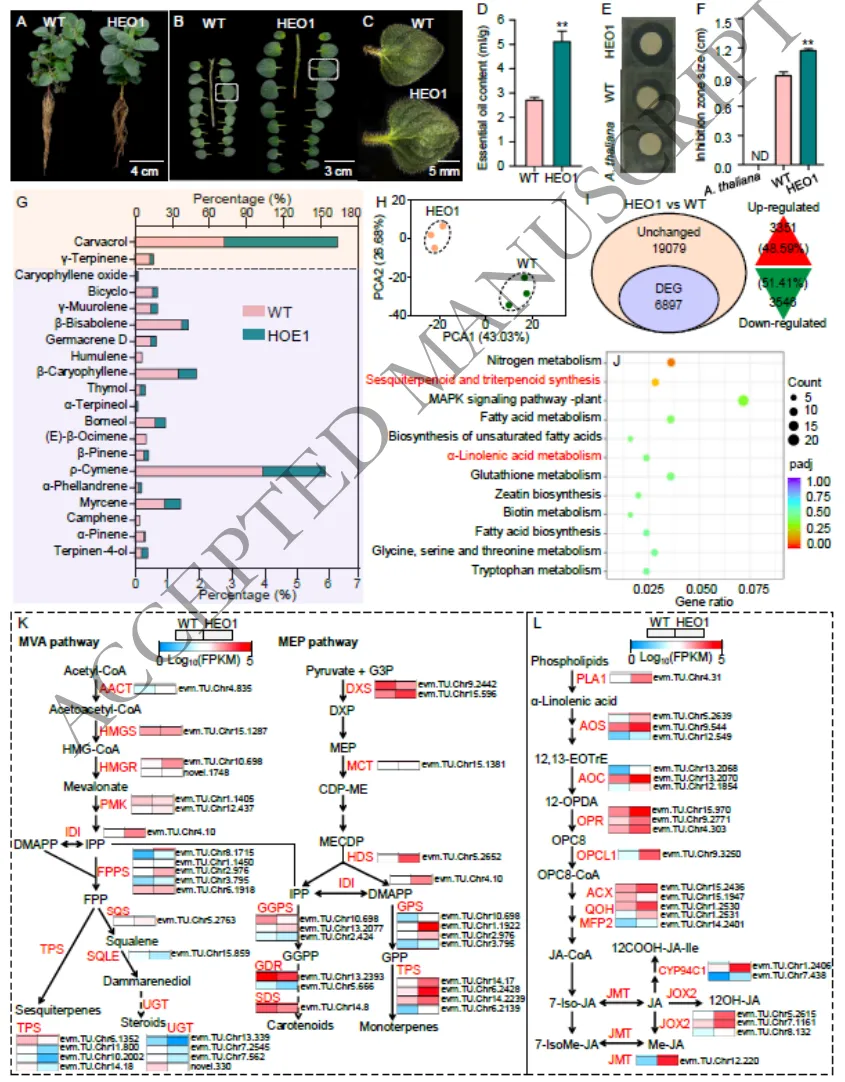
3.4 高精油含量希腊牛至突变体的筛选
快速筛选:利用精油对大肠杆菌的显著抑制作用,建立了一种高效的抑菌圈快速筛选体系,成功从诱变群体中初步筛选出51株候选突变体。
表型鉴定:鉴定出典型高精油突变体HEO1,其精油产量高达5.12%,比野生型高出88.24%,且表现出更大的叶片面积、厚度及更高的叶茎比。
成分解析:突变体HEO1的精油成分发生显著改变,香芹酚含量大幅提升,而其前体物质γ-松油烯含量降低,且染色体数目与野生型保持一致。
为克服传统精油提取耗时费力的瓶颈,基于牛至精油的抗菌特性开发了一种快速筛选体系,并成功从⁶⁰Co-γ射线诱导的突变体群体中筛选出51株具有更强抑菌活性的突变体。其中,代表性突变体HEO1的精油含量高达5.12%,较野生型(2.72%)显著提升了88.24%,并产生更大的抑菌圈。表型分析表明,HEO1具有更大的叶片尺寸、增加的叶片厚度以及更高的叶茎比。化学成分分析进一步证实,HEO1精油中香芹酚含量显著增加,而其前体γ-松油烯含量下降。细胞学观察确认该突变体未发生倍性改变,染色体数目维持在2n=30(Fig. 4A-G)(Fig. 1B)(Fig. S12-S15)。
图4.高精油突变体HEO1的表型、精油特征及比较转录组学分析。**图A-F展示了突变体HEO1与野生型在植株形态、叶片特征(面积、厚度、叶茎比)以及抑菌圈大小方面的直观对比;图G展示了HEO1与野生型在精油核心成分(如香芹酚、γ-松油烯等)含量上的显著性差异;图H-I展示了两者比较转录组学分析中差异表达基因(DEGs)的分布情况及qRT-PCR的验证结果;图J展示了差异表达基因的KEGG通路富集结果(如富集于萜类合成、α-亚麻酸代谢等);图K-L展示了MEP途径(单萜合成)及JA生物合成途径中关键基因的表达热图。
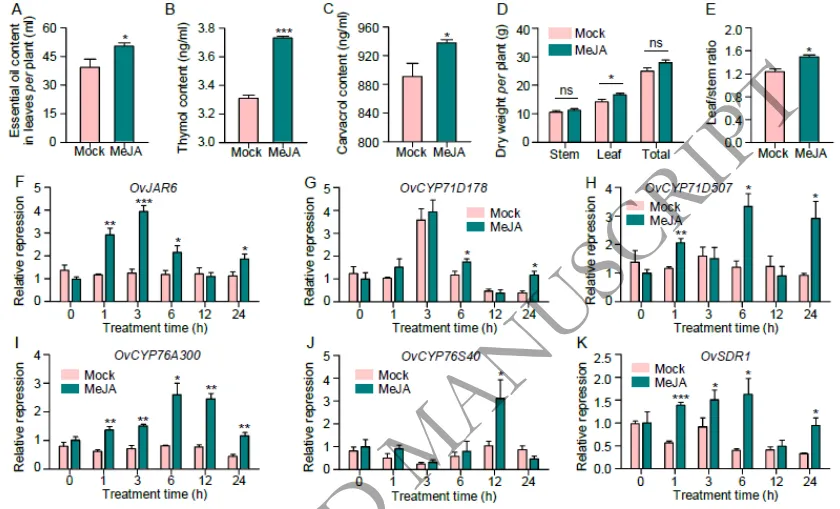
3.5 茉莉酸(JA)在希腊牛至精油合成中的作用
转录组学:比较转录组分析鉴定出6,897个差异表达基因,这些基因主要富集于萜类生物合成、α-亚麻酸代谢及JA生物合成等关键通路。
通路激活:突变体中MEP途径的单萜合成基因及JA生物合成途径的核心基因均呈现出显著的上调表达模式,与高精油表型高度吻合。
外源验证:外源喷施JA显著提升了野生型植株的精油产量及百里香酚、香芹酚含量,并同步上调了OvJAR6及OvSDR1等关键合成基因的表达。
通过对突变体HEO1与野生型植株进行比较转录组学分析,共鉴定出6,897个差异表达基因(DEGs)。通路富集分析表明,这些基因显著富集于倍半萜和三萜生物合成、α-亚麻酸代谢(JA生物合成途径)等网络。在分子层面上,MEP途径中负责单萜合成的11个基因中有10个在HEO1中显著上调,同时JA生物合成途径中的21个关键基因也呈现高表达,这与突变体中香芹酚和总精油含量的增加高度一致。为验证JA的调控功能,对外源喷施JA的植株进行检测,结果显示精油总产量增加了27.95%,百里香酚和香芹酚含量分别提升了12.69%和5.28%,且生物量指标显著改善。同时,JA处理强烈诱导了OvJAR6及OvSDR1、OvCYP71D178等精油合成关键基因的表达,充分证实了JA在正向调控牛至萜类生物合成中的核心作用(Fig. 4H-L)(Fig. 5)(Fig. S16)。
图5.外源茉莉酸(JA)处理对希腊牛至表型及基因表达的影响。图A-E展示了外源喷施JA处理后,野生型植株在精油总产量、核心成分(百里香酚和香芹酚)含量以及生物量指标上的显著提升;图F-K展示了JA处理后,精油合成关键基因(如OvJAR6、OvSDR1、OvCYP71D178等)表达量的显著上调情况。
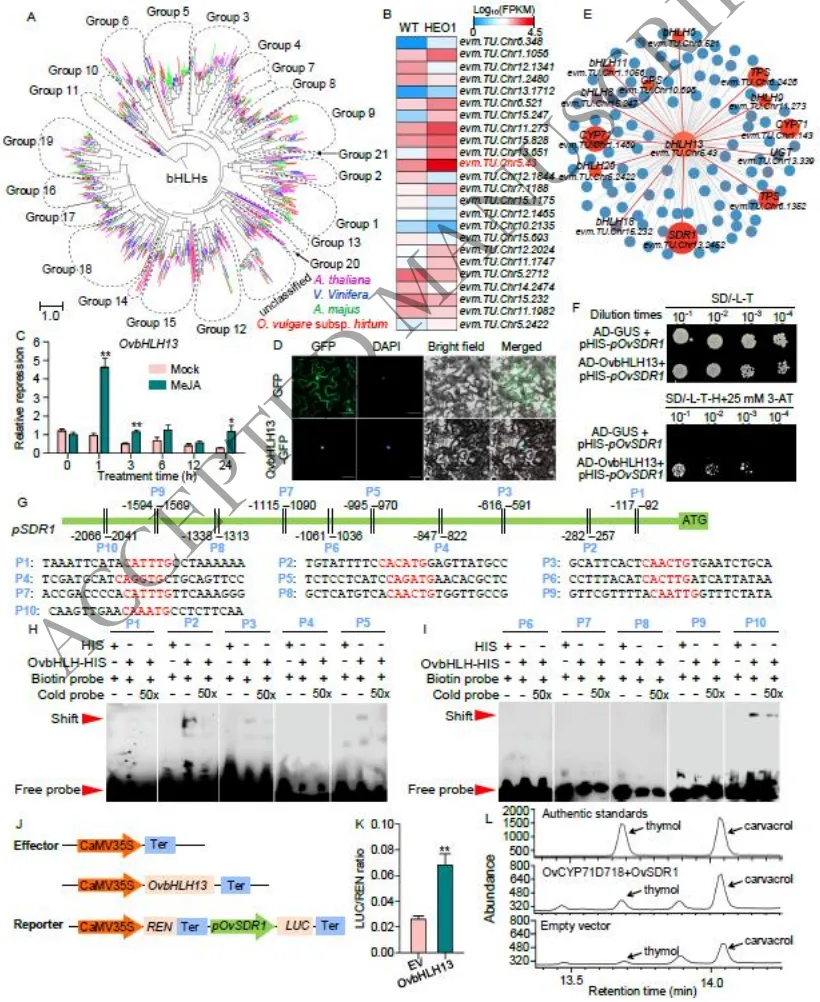
3.6 鉴定激活萜类合成基因OvSDR1表达的JA诱导型bHLH转录因子
因子挖掘:全基因组范围内鉴定出146个bHLH转录因子,其中OvbHLH13在突变体中表达量最高,且受外源JA强烈诱导,定位于细胞核。
靶标预测:共表达网络分析及启动子基序扫描揭示,OvbHLH13与核心合成基因OvSDR1高度共表达,且OvSDR1启动子区域含有多个E-box顺式作用元件。
功能确证:酵母单杂交、EMSA及双荧光素酶实验证实OvbHLH13直接结合并激活OvSDR1;烟草共表达实验进一步验证了该模块可显著提升百里香酚和香芹酚产量。
为解析JA介导的转录调控网络,在希腊牛至基因组中系统鉴定了146个bHLH转录因子。表达谱分析锁定了一个在HEO1突变体中表达量极高且受JA强烈诱导的候选基因OvbHLH13,亚细胞定位证实其编码蛋白特异性定位于细胞核。共表达网络分析显示,OvbHLH13与包括OvSDR1在内的多个萜类合成关键基因存在显著的共表达关系。序列分析发现OvSDR1启动子区域包含10个潜在的bHLH结合位点(E-box)。随后,通过酵母单杂交(Y1H)、凝胶阻滞实验(EMSA)以及植物体内双荧光素酶(Dual-LUC)反式激活实验,多维度确证了OvbHLH13能够直接结合OvSDR1启动子并强效激活其转录。最后,在本氏烟草中瞬时共表达OvCYP71D178与OvSDR1,显著促进了百里香酚和香芹酚的生物合成,从而完整揭示了“OvbHLH13-OvSDR1”这一控制希腊牛至精油合成的核心调控模块(Fig. 6)。
图6.转录因子OvbHLH13的鉴定及其对OvSDR1的转录激活功能验证。图A-B展示了bHLH转录因子家族的系统发育分析及在突变体中的差异表达情况(锁定高表达的OvbHLH13);图C-D显示了OvbHLH13受外源JA强烈诱导的表达特性及其在细胞核中的特异性定位;图E揭示了OvbHLH13与核心合成基因(如OvSDR1)的共表达网络;图F-K通过酵母单杂交(Y1H)、凝胶阻滞实验(EMSA)及双荧光素酶(Dual-LUC)反式激活实验,多维度证实了OvbHLH13对OvSDR1启动子的直接结合与强效激活作用。






 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
