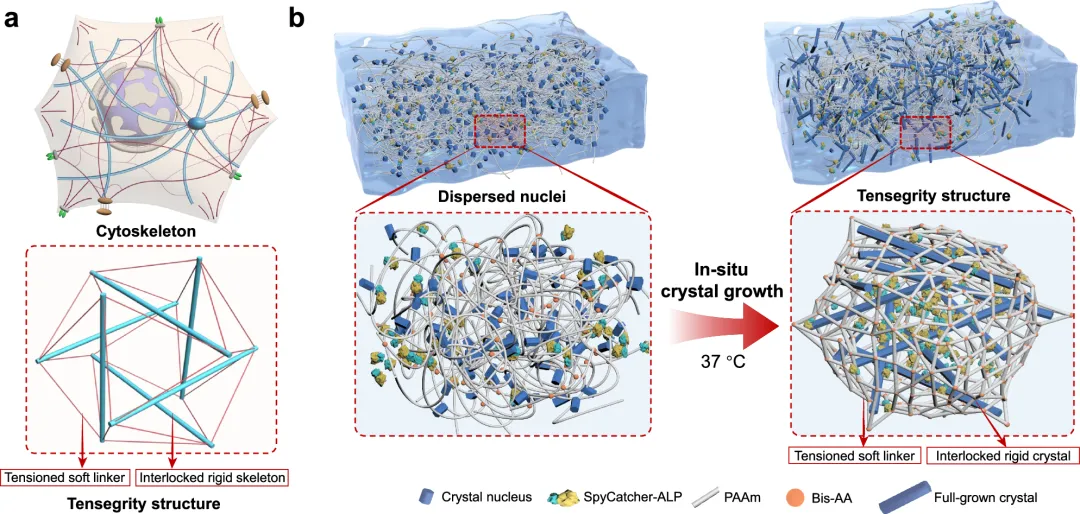
张力完整性结构是由孤立的刚性压缩部件构成的,这些部件通过连续的拉伸部件网络固定在适当位置,它们在诸如细胞外基质和细胞骨架等自然系统中起着核心作用。这些结构具备动态生物功能所必需的非对称机械特性。在此,我们介绍了一种合成方法,用于设计具有张力完整性结构的水凝胶,其灵感来源于生物系统背后的力学化学原理。通过在预先形成的聚合物网络中利用酶诱导的氨基酸晶体生长,我们实现了微晶体棒在预受力聚合物基质中随机互锁的分层集成。这种设计模仿了自然的张力完整性结构,能够平衡机械力以保持高刚度(拉伸模量高达 30 兆帕)、断裂韧性(2600 焦耳/米²)和含水量(超过 80%)。所得的水凝胶由于其张力完整性结构而表现出双模量特性,其拉伸与压缩模量之比为 13。这种仿生学方法为开发用于组织工程及其他领域的坚固且具有自适应性的材料提供了一种策略。
水凝胶一直扮演着独特而矛盾的角色。它由亲水高分子网络构成,能锁住大量水分,模拟生物组织的柔软湿润环境,是组织工程、柔性电子等领域的理想候选材料。然而,传统水凝胶的力学性能往往堪忧——“软”是其固有标签,高含水量通常伴随着低强度、低韧性的代价。长期以来,科学家们努力模仿自然界生物的智慧,试图让水凝胶“刚柔并济”。细胞骨架和人体筋膜系统为我们提供了绝佳蓝图:它们都基于“张拉整体”结构——离散的刚性支柱(如微管、骨骼)被连续的张力网络(如肌动蛋白、肌腱)所包裹和拉紧,这种结构实现了高效的力量传递、高比强度,并展现出独特的非互易力学行为(例如,软骨在拉伸和压缩时硬度截然不同)。然而,在合成材料中,尤其是水凝胶里,精准复现这种力化学耦合的精妙结构,在保持超高水含量的同时,赋予其高强度、高韧性和显著的双模量特性,一直是一个难以逾越的挑战。核心技术:在凝胶内部“种”出钢筋网络
性能飞跃:数据背后的“力学奇迹”
超高强韧:拉伸模量最高可达30 MPa,断裂能高达2600 J m⁻²,远超许多已报道的高强水凝胶,甚至媲美天然软骨。
高水含量:在获得如此卓越力学性能的同时,含水量依然保持在80%以上,没有牺牲其生物相容性的基础。
核心特性——双模量:其拉伸模量是压缩模量的13倍!这种显著的“非互易”力学行为,使其能像真正的生物组织一样,对拉伸和压缩做出截然不同的响应。
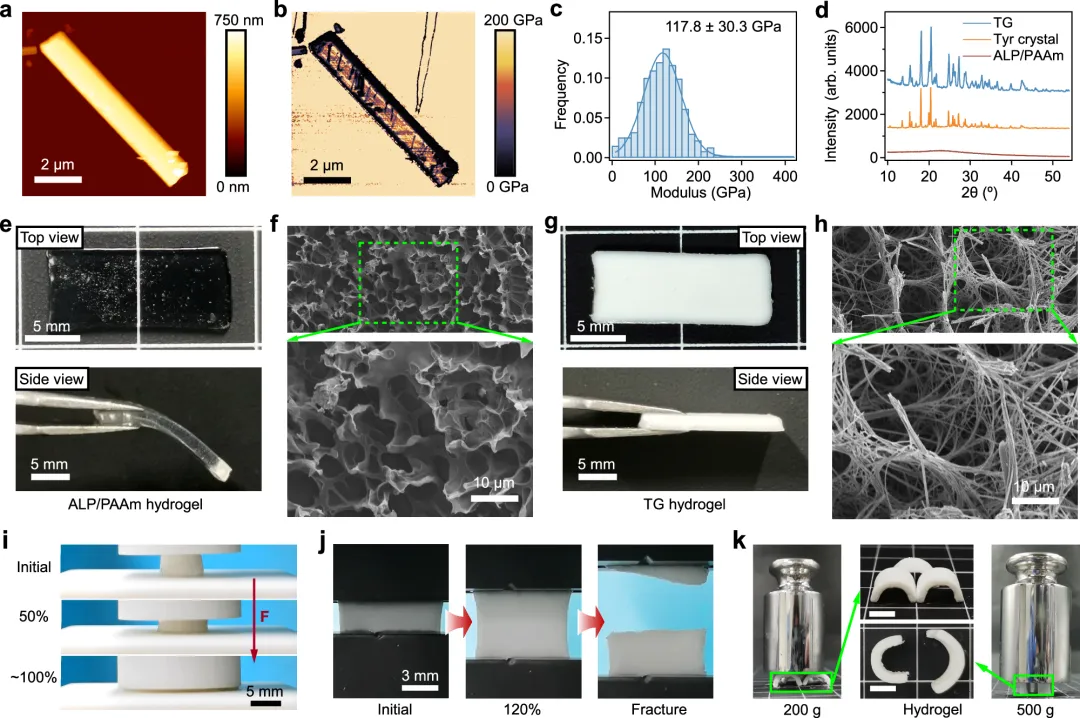
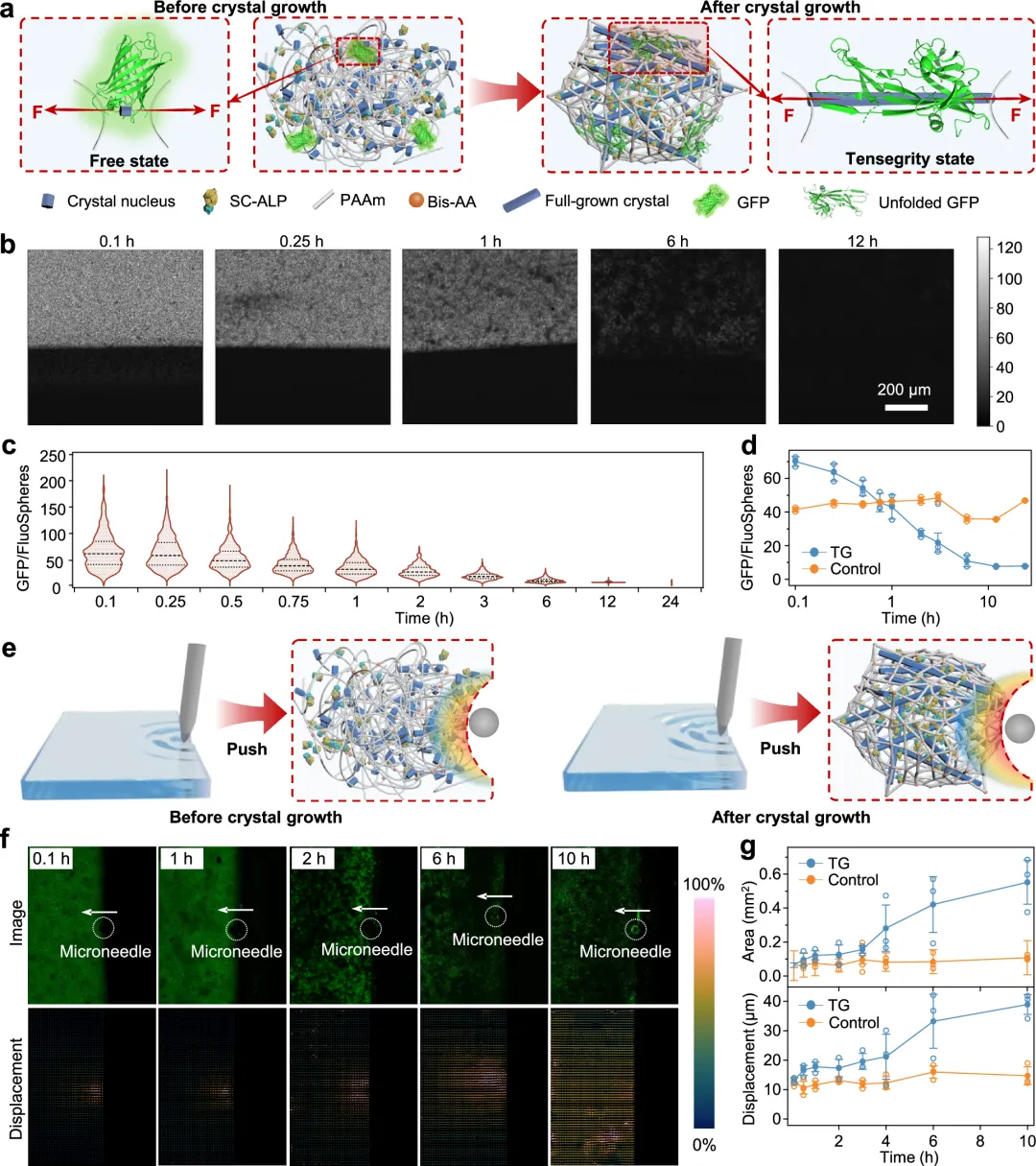
图1 通过氨基酸晶体棒的原位生长设计水凝胶张力整合结构。由微管和肌动蛋白丝形成的细胞骨架图示,以及由棍子和绳索形成的张力整合结构(下图)。b 通过氨基酸晶体棒的原位生长,在水凝胶中张力整合结构的形成示意图。ALP被共价整合进水凝胶网络,以诱导氨基酸晶体棒的原位生长。晶体棒对弹性网络施加较大张力,而网络对晶体棒施加压缩,形成张力整合结构。图2 酪氨酸晶体和张力整合结构水凝胶(TG水凝胶)的表征。a, b 基于AFM纳米凹陷的酪氨酸单晶典型拓扑高度(a)和模量图(b)。c 酪氨酸单晶的统计模量分布。d 粉末酪氨酸晶体(Tyr晶体)、TG和ALP/PAAm水凝胶的XRD图谱。e、f ALP/PAAm水凝胶的光学(e)和SEM(f)图像。g, h TG水凝胶的光学(g)和SEM(h)图像。厚网格状结构由晶体棒状结构形成,如SEM图像所示。i 初始和压缩TG水凝胶的图像(应变 ~ 50% 和 ~ 100%)。虚线框表示TG水凝胶的初始形状。j 初始、拉伸(应变 ~120%)及断裂TG水凝胶的图像。k 倾斜TG水凝胶,负载不同重量。三块带状水凝胶带状面可承载200克,带弧度面可承载200克,两条带状带状水凝胶侧面可承重500克。中间图像中的比例条代表5毫米。图3 TG水凝胶聚合物网络中的张力。示意图展示了使用绿色荧光蛋白(GFP)作为荧光力探头探测内部张力的实验装置。GFP由晶体生长驱动的聚合物拉伸展开,导致荧光减弱。b 同一TG水凝胶中不同时间晶体原位生长后GFP/FluoSpheres比率图。c TG水凝胶在原位晶体生长后不同时间(0.1、0.25、0.5、0.75、1、2、3、6、12和24小时)GFP/FluoSpheres比值总结。虚线表示平均值(n = 1107)。d TG和对照(ALP/PAAm)水凝胶在原位晶体生长后不同时间的平均GFP/FluoSpheres比值。e 示意图展示了在施加应变下TG水凝胶变形场的实验装置。绿色荧光微球固定在水凝胶网络中,使用微针记录晶体生长前后在施加应变(~ 170微米)下生长前后,并进行分析。f 在同一TG水凝胶中,不同时间(0.1、1、2、6和10小时)晶体生长后,微球的荧光图像(图像)和形态场(位移)。变形场中的小箭头表示微球位移的方向和距离。右侧的颜色条表示微球位移距离,最高位移设为100%。g TG中的变形场面积(上方)和微球位移面积(下方)汇总,控制水凝胶在不同时间(0.1、0.5、1、2、3、4、6和10小时)晶体生长后。无晶体生长的ALP/PAAm水凝胶作为对照。(d和g)中的值代表平均和标准差(n = 3)。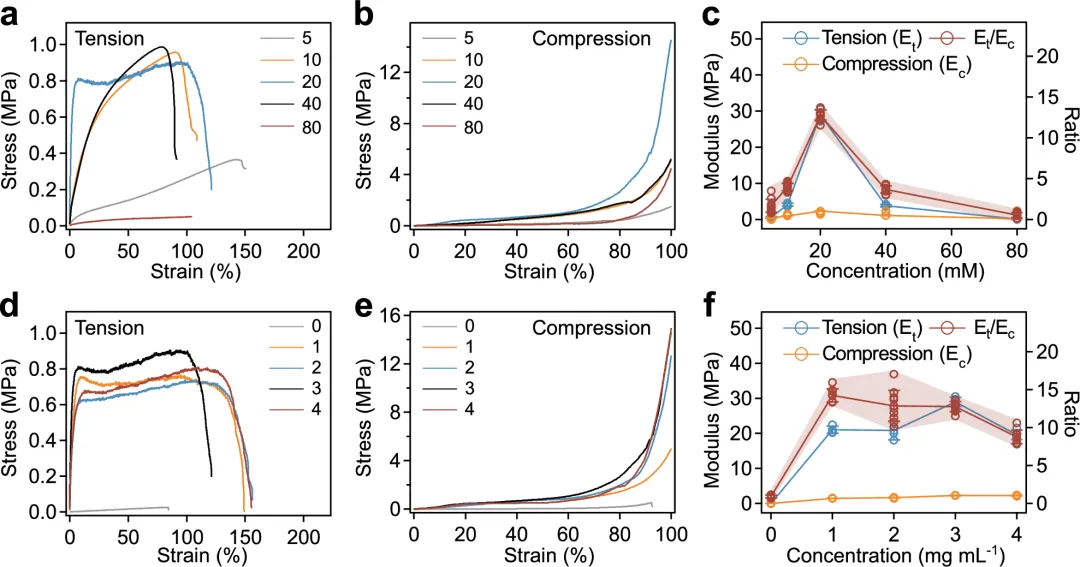
图4 张力整合结构水凝胶的力学性能。a, b TG水凝胶在不同浓度双丙烯酰胺(5–80 mM)下,在拉伸(a)和压缩(b)下的典型应力-应变曲线。晶体核和SC-ALP的浓度分别为2和3 mg mL−1,分别。c 总结了拉伸模量和压缩杨氏模量,以及拉伸模量与压缩模量的比值(Et/Ec)在不同浓度的双丙烯酰胺(5–80 mM)下对TG水凝胶的分析。数值表示平均差和标准差(E 为 n = 3)。t以及Ec; 对于E,n = 9t/Ec). d, e TG水凝胶在不同SC-ALP(0–4 mg mL)浓度下,在拉伸(d)和压缩(e)下的典型应力-应变曲线−1). 双丙烯酰胺和晶核的浓度分别为20 mM和2 mg mL−1,分别。f 拉伸模和压缩杨氏模量的总和,以及拉伸模量与压缩模量的比值(Et/Ec不同浓度的 SC-ALP(0–4 mg mL)中 TG 水凝胶的−1). 值表示平均差和标准差(E 的 n = 3t以及Ec; 对于E,n = 9t/Ec).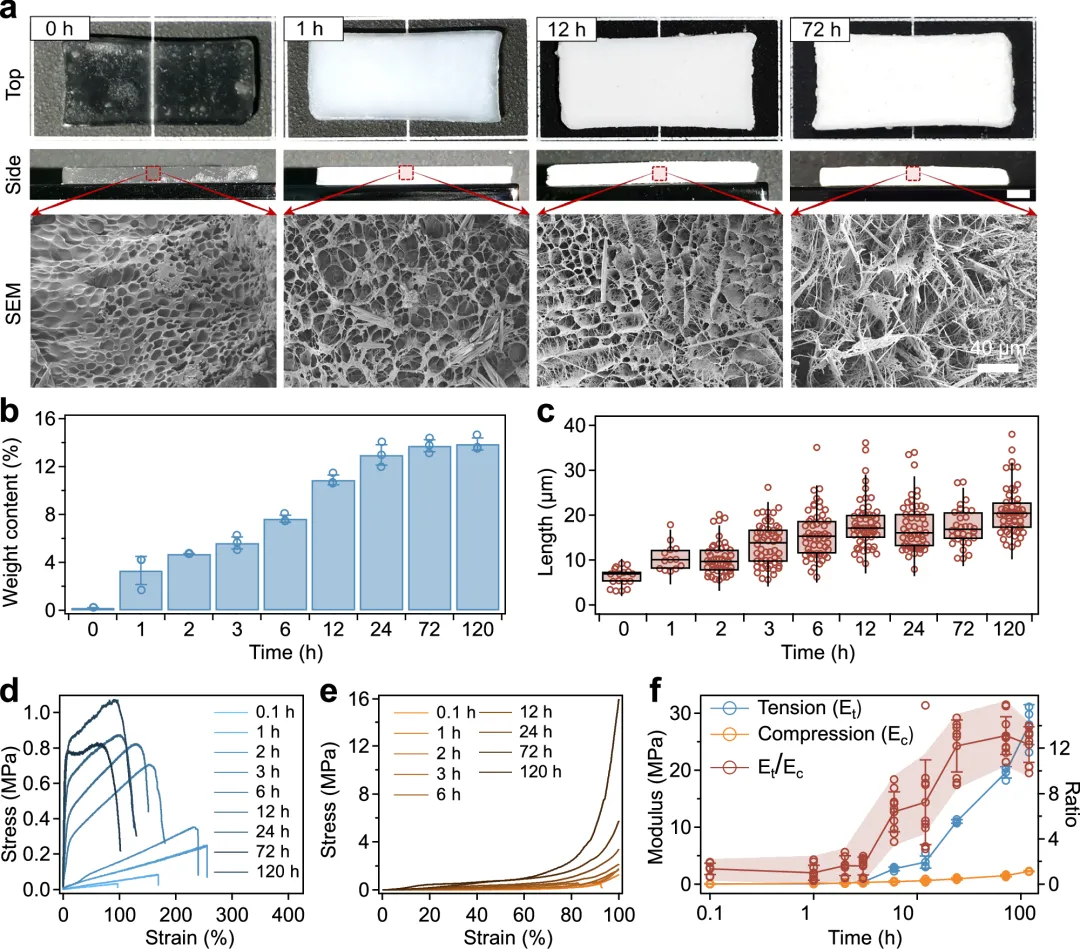
图5 张力整合结构形成的动力学。一组在不同时间(0、1、12和72小时)原位晶体生长后,带状的TG水凝胶的光学(上方)和SEM(下方)图像。光学图像的比例条为5毫米b,c,TG水凝胶中晶体在不同时间的原位晶体生长后,其重量含量(b)和长度(c)。(b)中的值表示均值和标准差(n = 3)。(c)中的中间线代表中位数,胡须表示标准差(n = 17、13、46、54、54、54、54、54、54、54、30和54,分别对应0、1、2、3、6、12、24、72和120小时)。d, e TG水凝胶在原位晶体生长后,在拉伸(d)和压缩(e)下的典型应力-应变曲线,适用于不同时间(0.1、1、2、3、6、12、24、72和120小时)。f 总结拉伸模量与压缩模量 杨氏模量及拉伸模量与压缩模量比值(Et/EcTG水凝胶在原位晶体生长后,在不同时间(0.1、1、2、3、6、12、24、72和120小时)中获得。值表示平均差和标准差(E 的 n = 3t以及Ec; 对于E,n = 9t/Ec).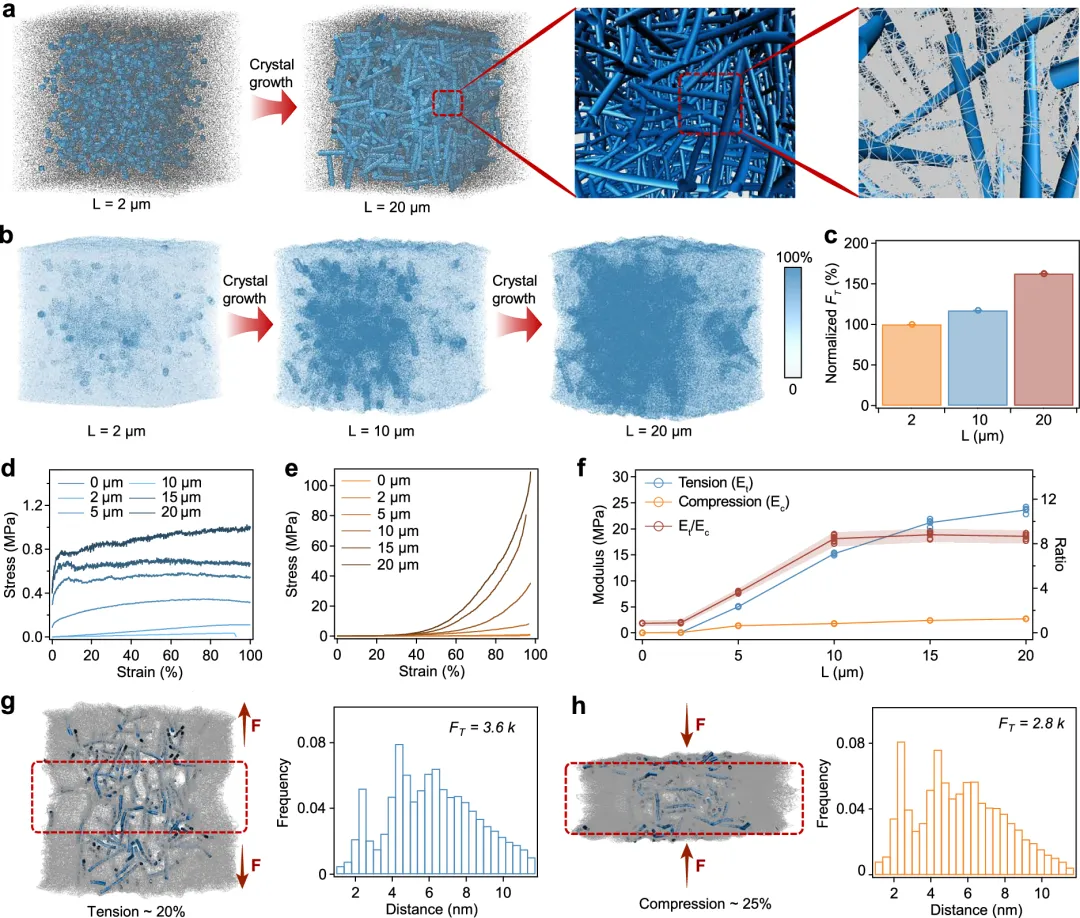 图6 张力整合结构和TG水凝胶力学性能的分子动力学模拟。A 水凝胶内部晶体生长和张力整合结构的快照。最右侧快照中的白线表示晶体棒周围的预应力聚合物网络。b 晶体生长过程中聚合物网络上的张力快照。晶体棒的长度分别为2、10和20微米。右侧的颜色条表示TG水凝胶聚合物网络上的归一化拉伸力,最高拉伸力设为100%。c 归一化平均拉伸力(FT随着晶体棒的生长,聚合物网络上的数量增加。在2微米晶体棒长处,平均拉伸力设为100%。数值表示均值和标准差(n = 3)。d, e TG水凝胶在不同晶体长度(0、2、5、10、15和20微米)下,在拉伸(d)和压缩(e)下的典型应力-应变曲线。f 总结拉伸模量和压缩模量,以及拉伸模量与压缩模量的比值(Et/Ec)在不同晶体长度(0、2、5、10、15和20微米)的TG水凝胶中。值表示平均差和标准差(E 的 n = 3t以及Ec;对于E,n = 9t/Ec).g 在20%拉伸应变下,张力整合结构(左)及相邻交联(右)间的统计距离快照。每个交联位点的平均拉伸力(FT)为3.6k,其中k对应于聚丙烯酰胺链的弹簧常数。h 张力整合结构快照(左)及水凝胶网络中相邻交联点之间的统计距离,在25%压缩应变下。每个交联位点的平均拉伸力(FT)为2.8k,其中k对应聚丙烯酰胺链的弹簧常数。
图6 张力整合结构和TG水凝胶力学性能的分子动力学模拟。A 水凝胶内部晶体生长和张力整合结构的快照。最右侧快照中的白线表示晶体棒周围的预应力聚合物网络。b 晶体生长过程中聚合物网络上的张力快照。晶体棒的长度分别为2、10和20微米。右侧的颜色条表示TG水凝胶聚合物网络上的归一化拉伸力,最高拉伸力设为100%。c 归一化平均拉伸力(FT随着晶体棒的生长,聚合物网络上的数量增加。在2微米晶体棒长处,平均拉伸力设为100%。数值表示均值和标准差(n = 3)。d, e TG水凝胶在不同晶体长度(0、2、5、10、15和20微米)下,在拉伸(d)和压缩(e)下的典型应力-应变曲线。f 总结拉伸模量和压缩模量,以及拉伸模量与压缩模量的比值(Et/Ec)在不同晶体长度(0、2、5、10、15和20微米)的TG水凝胶中。值表示平均差和标准差(E 的 n = 3t以及Ec;对于E,n = 9t/Ec).g 在20%拉伸应变下,张力整合结构(左)及相邻交联(右)间的统计距离快照。每个交联位点的平均拉伸力(FT)为3.6k,其中k对应于聚丙烯酰胺链的弹簧常数。h 张力整合结构快照(左)及水凝胶网络中相邻交联点之间的统计距离,在25%压缩应变下。每个交联位点的平均拉伸力(FT)为2.8k,其中k对应聚丙烯酰胺链的弹簧常数。我们这种张力结构水凝胶的机械性能突显了机械化学在材料设计中的关键作用。通过在预受力的聚合物网络中利用酶诱导的氨基酸结晶,我们展示了一种将化学合成与机械工程巧妙结合的方法,以实现刚度、韧性以及高保水性的平衡。原位晶体生长过程通过内部结构演变自然地引发机械性能的显著转变,而非通过外部施加的各向异性实现。这种方法受到了生物系统(如细胞外基质和细胞骨架)的启发,在这些系统中,化学相互作用与机械力之间的协调导致了适应性强、有弹性的结构。
我们的方法超越了传统的水凝胶工程,着重于机械化学过程的协同效应。尽管去磷酸化及其后续晶体生长的化学过程此前已有相关报道,但本研究中在位生长的氨基酸晶体棒将刚性和柔性的成分整合到一个坚固的结构中,从而形成了张力结构体系。这一过程不仅在聚合物网络中引入了预应力条件,还能够形成相互连接的晶体单元,这些单元在不牺牲水含量的情况下增强了水凝胶的结构,从而实现了具有独特机械性能的体系。这种结构设计与机械化学相互作用的特殊结合,使得水凝胶具备了一种在缺乏张力结构体系的水凝胶中无法实现的机械性能组合。这种机械化学相互作用对于复制自然组织中观察到的非对称模量特性至关重要,在这些组织中,在拉伸和压缩条件下会表现出不同的机械反应。
实验证据,包括扫描电子显微镜(SEM)和原子力显微镜(AFM)图像,清晰地表明了在水凝胶内部形成了相互嵌套的晶体结构;而基于荧光的检测则直接表明了由于晶体生长而产生的内部张力。张力架结构的存在有助于解释所观察到的机械行为,包括在拉伸和压缩测试中高达 13 的拉伸与压缩模量比值。分子动力学模拟进一步说明了机械化学如何驱动这些张力架结构的形成。模拟显示,在晶体生长过程中聚合物网络内产生的张力会传播远距离应力,增强了机械完整性,并使材料具有双模量响应。这种响应对于这些水凝胶的特性至关重要,使它们能够模拟生物组织(如软骨)中的机械不对称性。
与传统的纤维增强或双网络设计等方法相比,我们的方法将机械化学控制与分层自组装相结合,从而制备出具有增强性能的水凝胶。预应力聚合物与原位生长的晶体的结合为在机械加载时立即进行应力传递提供了有效手段,即使在显著的应变条件下也能保持结构的完整性。
要在软性材料内部构建一种张力-压缩结构,需要从多个关键方面进行合理设计。首先,需要设计具有不同刚度的元件,其中刚性元件充当相互连接的刚性骨架,而柔性强的元件则作为柔性的连接件。其次,具有大长径比的刚性元件应具有无序的排列方式,以便在低质量含量的情况下实现机械连接。第三,柔性的连接件或弹性柔性的网络必须进行预应力处理,以便在机械加载时能够立即进行应力传递,即使在显著的应变条件下也能保持结构的完整性。遵循这些规则,这种张力-压缩结构也可以扩展到其他水凝胶系统中。例如,通过利用水热过程或催化还原在大长径比下诱导无机盐的原位晶体生长,可以在不进行生物合成步骤的情况下在水凝胶中实现张力-压缩结构。此外,张力-压缩结构也可以在其他软性材料(如干燥弹性体)中实现。例如,通过在软聚合物网络中引入一种含有刚性、坚硬且各向异性聚合物前体的物质,当在软聚合物网络中施加拉伸力并触发刚性聚合物的交联时,刚性聚合物所产生的张力能够引发张力结构体的形成。
这些经过设计的张力结构体系具有独特的机械优势,包括高强度与轻质的比值以及非对称的机械响应——这些特性是生物组织所具备的基本特征。受到此启发,结合张力结构的生物启发型水凝胶设计为开发用于生物医学和仿生应用的坚固且具有自适应性的材料提供了一种有前景的策略。例如,张力结构水凝胶可以作为高含水量的材料。
关注【材料探界新语】,带你第一时间探访材料科学前沿!原文:https://doi.org/10.1038/s41467-025-58956-3