Plant Physiol|南京农大唐设研究组发现高温加速水稻叶片衰老的同时会促进氮素运输并提高籽粒蛋白合成
- 2026-06-18 11:14:10
特别提醒,本文的图片序号与原文不一致
水稻是供给全球20%膳食能量的主粮,稻米优质蛋白是膳食关键营养来源,但IPCC预测本世纪末全球地表温度将提升1.0–5.7℃,灌浆期高温成为制约水稻稳产提质的全球性胁迫因素。高温会破坏花药与花粉发育降低结实率、缩短灌浆周期、减少粒重,同时紊乱淀粉与蛋白合成代谢,劣化加工与食味品质;籽粒70%–90%氮素来自花前营养体再转运,叶片是籽粒氮最主要源器官(贡献60%再转运氮),谷氨酰胺、天冬酰胺是韧皮部氮长距离转运核心形态,GS-GOGAT循环、OsLHT1/OsAAP6转运蛋白共同主导氮素的同化与跨器官运输。叶片衰老是源氮再分配的关键生理过程,逆境诱导的早衰可重塑源库关系,但升温如何通过调控叶片衰老、氮转运差异化改变强弱粒蛋白储量,以及增氮与高温互作调控籽粒营养积累的分子代谢逻辑,既往研究缺少强弱穗分粒系统性解析,尤其代谢层面关键标志物尚未明确,基于此本研究通过大田原位增温 + 氮素调控 + 生理 + 代谢组多维度试验,填补升温下水稻强弱粒氮分配机理的研究空白。
2026年6月2日,南京农业大学农学院唐设研究组与安徽省农科院水稻所习敏研究组合作,在Plant Physiology杂志发表题为Elevated temperature accelerates leaf senescence and promotes leaf nitrogen transport to improve rice grain protein synthesis的研究论文。本研究依托大田开放式增温设施(白天升温2.28℃、夜间升温4.33℃),设置常规对照CK、灌浆期增温ET、常温增氮CKN、升温配施氮肥ET(额外60 kg/hm2氮素)四大处理,系统区分强势粒(SS,上部一次枝梗、灌浆快粒重高)与弱势粒(IS,二次枝梗、开花滞后灌浆迟缓)开展生理与代谢组联合测定。结果显示升温使强势粒千重下降2.1%,弱势粒粒重提升4.23%、蛋白含量上升2.95%;灌浆9–15 d升温诱导叶片游离氨基酸富集、上调GS/GOGAT关键氮同化酶活性与氨基酸转运基因OsLHT1、OsAAP6表达,促使20 d花后叶片程序性早衰,加速源叶氮素向籽粒转运;无论常温还是升温环境,外源补氮均可延缓叶片衰老、同步提升强弱穗籽粒蛋白积累。非靶向代谢组证实升温加速叶片氨基酸耗竭,L-组氨酸、D-棉子糖、龙胆酸三类关键胁迫代谢物富集是弱势粒蛋白提升的重要物质基础,该成果系统阐明气候变暖背景下水稻籽粒蛋白差异化积累的源库调控通路,为暖区水稻提质栽培提供理论依据。
本研究以热敏感粳稻武育粳3号为试验材料,依托FATE开放式田间红外增温系统精准模拟昼夜差异化升温(自然昼/夜温24.8℃/19.54℃,增温后27.08℃/23.87℃),设置四类栽培处理,从产量性状、叶片光合生理、活性氧代谢、源氮转运生理、籽粒蛋白动态、非靶向代谢组六大维度开展系统研究。
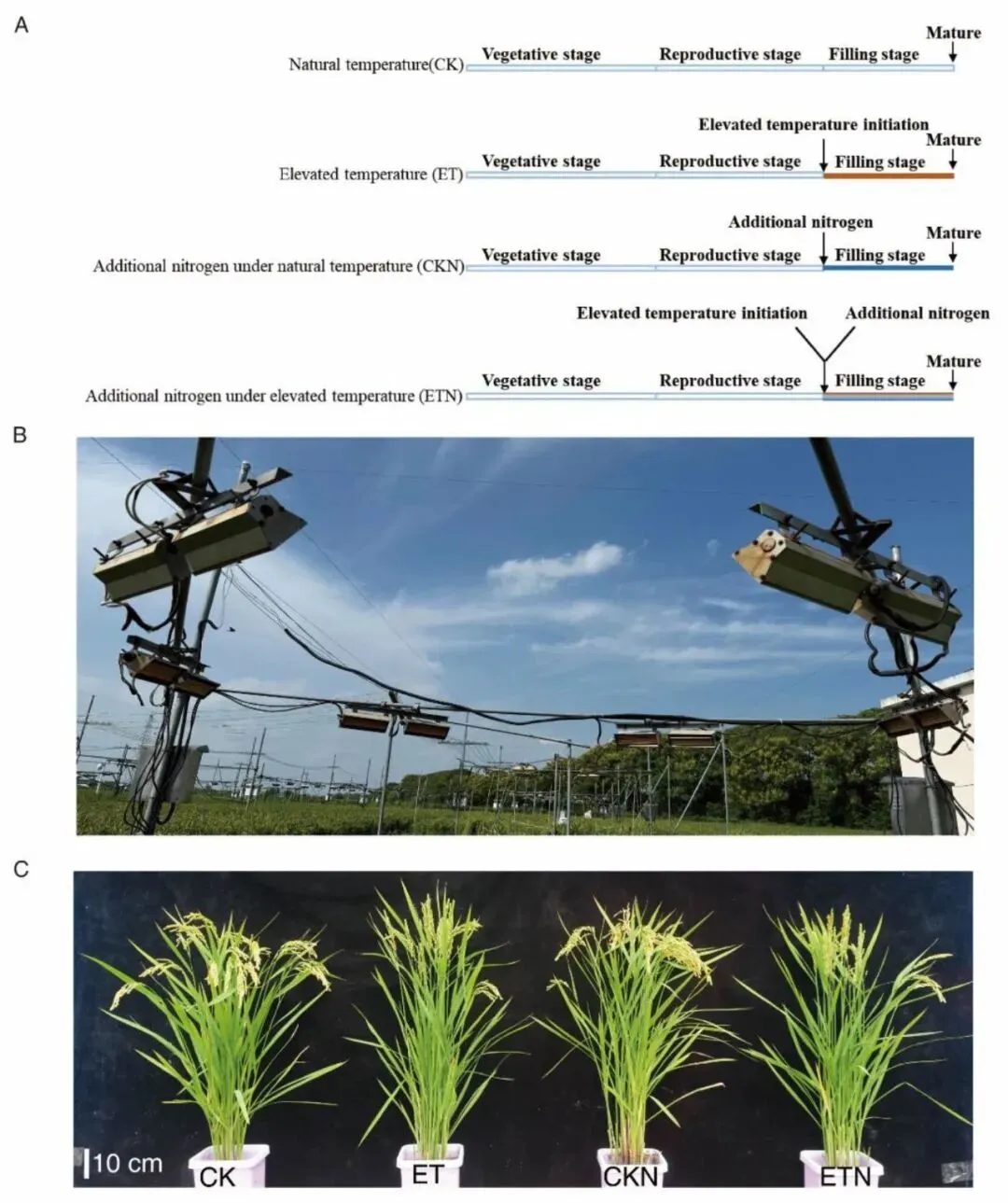
图1.田间增温试验设计示意图、FATE红外加温田间实景、花后30 d各处理水稻田间整株表型照片。
在产量构成层面,单纯升温ET造成结实率下降16.08%、千重降低3.05%、总产量降幅18.98%;常温补氮CKN千重下降14.42%、减产6.77%;升温叠加补氮ETN减产幅度最大,达18.98%,表明升温与过量后期施氮均不利于水稻干物质积累,二者叠加会进一步加剧产量损失,根源在于氮代谢增强打破植株碳氮平衡,碳水化物供给不足限制籽粒库容扩充。
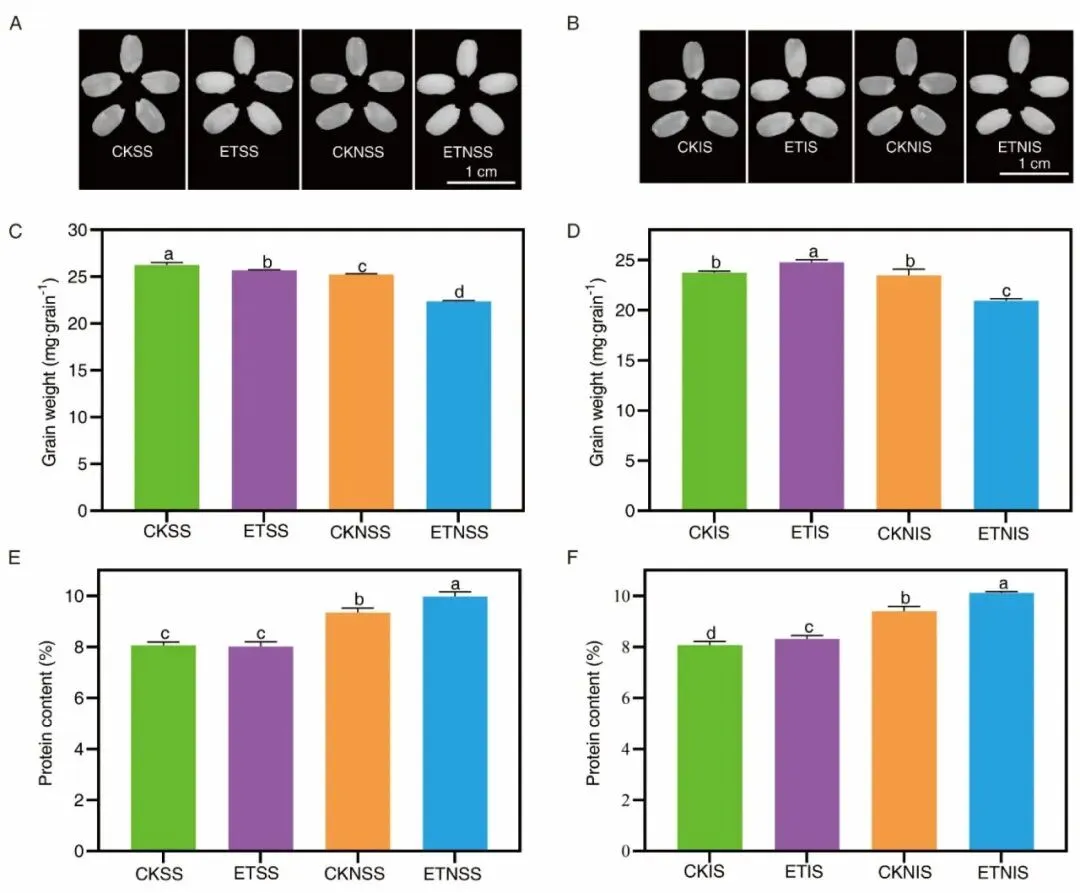
图2.成熟期强弱糙米扫描形态、千粒重与籽粒总蛋白含量统计结果。
叶片光合与衰老生理结果表明,灌浆进程中旗叶、倒二叶、倒三叶SPAD值持续走低,ET处理花后20–45 d倒二叶SPAD显著下降,30 d时旗叶尖端出现明显黄化早衰表型;升温在灌浆早期(6、9 DAF)短暂提升净光合速率Pn,但12 DAF后光合持续受抑,Fv/Fm 光化学效率从9 DAF开始显著降低,标志叶片遭遇持续性高温胁迫。高温诱导叶片H2O2大量累积(9 DAF增幅 28.41%、15 DAF增幅61.47%),同步上调APX、CAT、POD三类抗氧化酶活性以清除活性氧;而外源补氮无论常温或升温条件均可显著下调H2O2含量、提升POD活性,通过维持ROS稳态延缓叶片衰老,从生理层面解释氮肥缓解高温损伤的机理,但补氮无法完全抵消高温对光合系统的不可逆破坏。
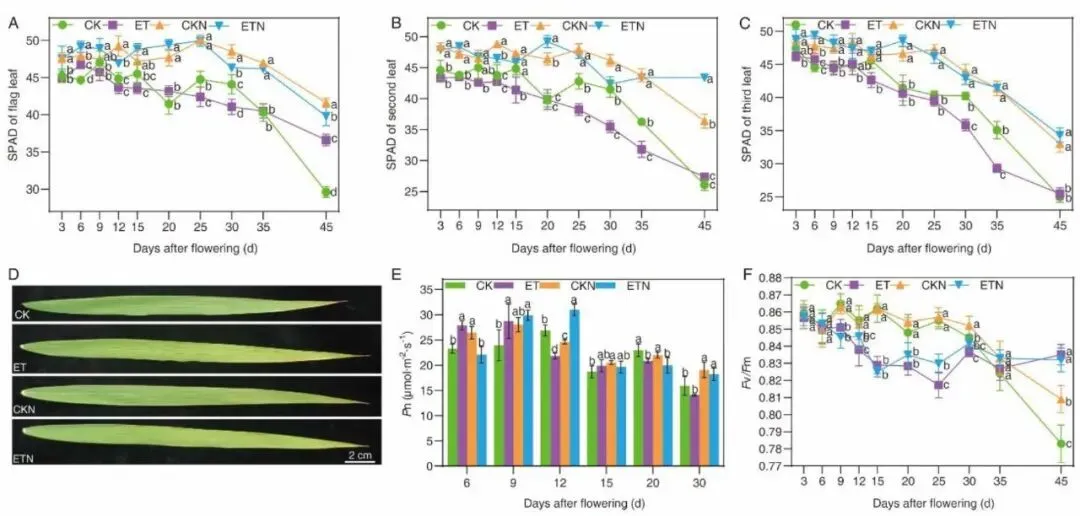
图3.灌浆期升温与补氮对水稻旗叶、倒二、倒三叶SPAD、净光合Pn、PSII最大光化学效率Fv/Fm及旗叶田间表型的影响。
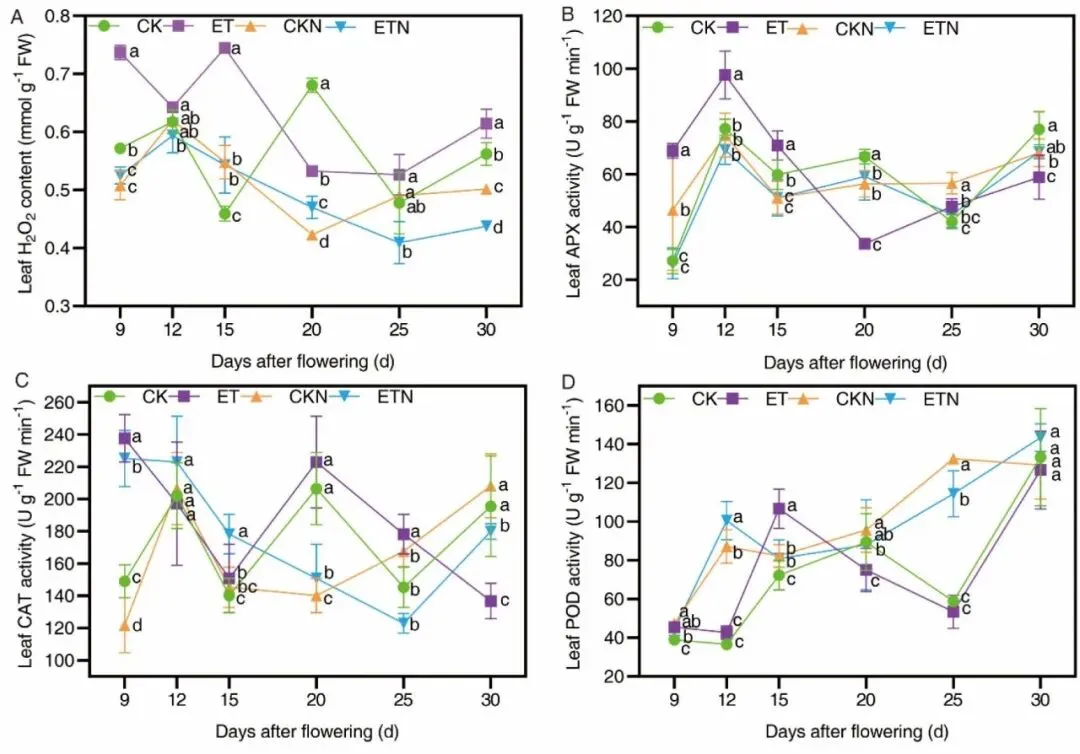
图4.不同处理叶片H2O2含量与APX、CAT、POD三种抗氧化酶活性动态变化。
源器官氮转运生理数据显示,ET处理20–45DAF 叶片全氮含量显著下降,9–15 DAF升温诱导叶片可溶性蛋白、游离氨基酸富集,GS、GOGAT氮同化酶活性大幅提升(GS 9/15 DAF分别 + 29.44%、+33.68%;GOGAT 12/15 DAF分别+ 91.33%、+128.41%),OsLHT1、OsAAP6氨基酸转运基因在9–20 DAF特异性上调,驱动叶片氮素快速拆解并以氨基酸形态经由茎鞘向籽粒转运,最终造成20 DAF后叶片氮匮乏、启动加速衰老程序;补氮处理全生育期维持叶片更高氮与游离氨基酸水平,GOGAT活性持续高位,转运基因表达进一步增强,在延缓衰老的同时持续为籽粒供给氮源。
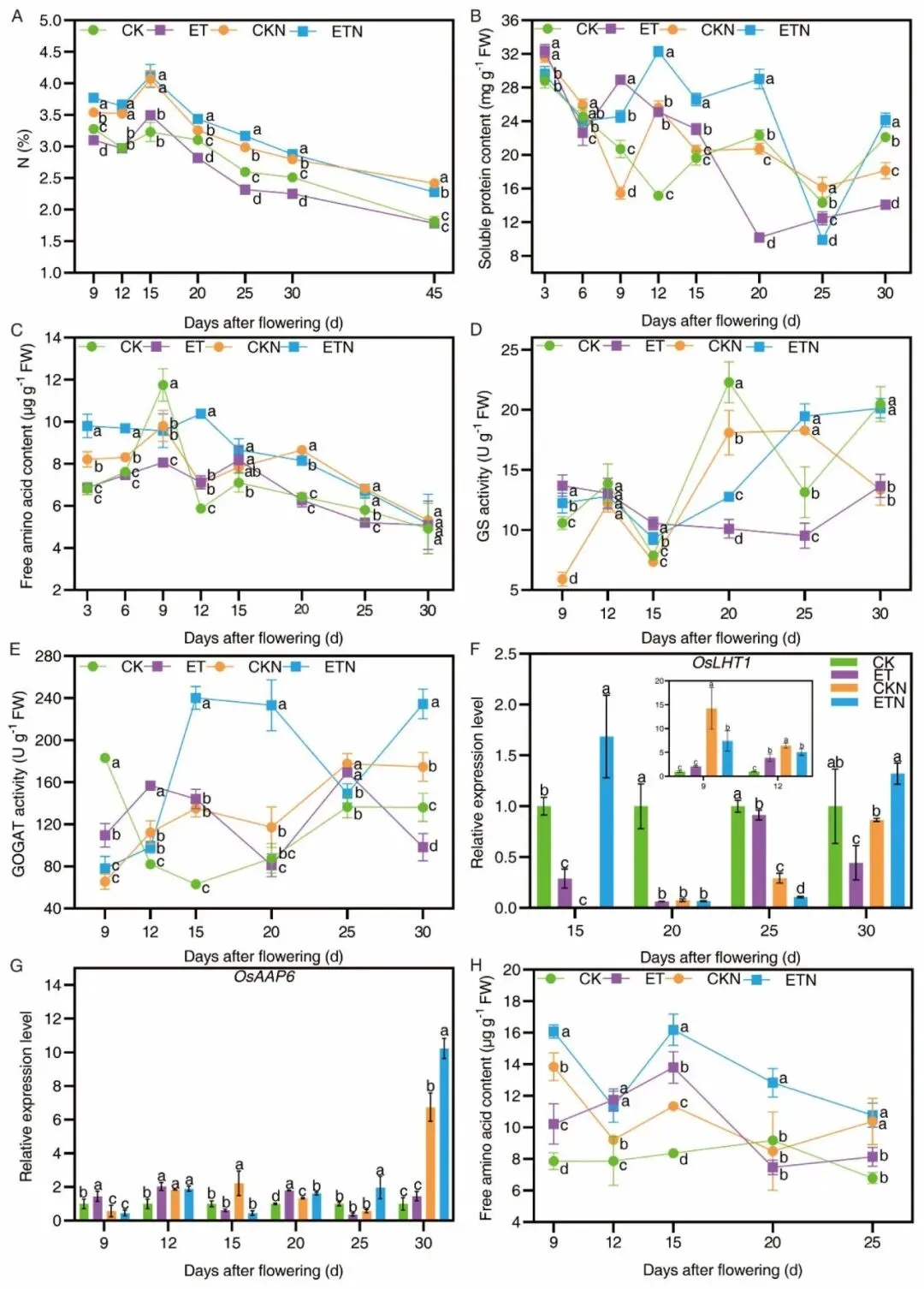
图5.表征温氮处理调控叶片全氮、可溶性蛋白、游离氨基酸、GS/GOGAT酶活、OsLHT1/OsAAP6基因表达以及茎鞘游离氨基酸积累规律。
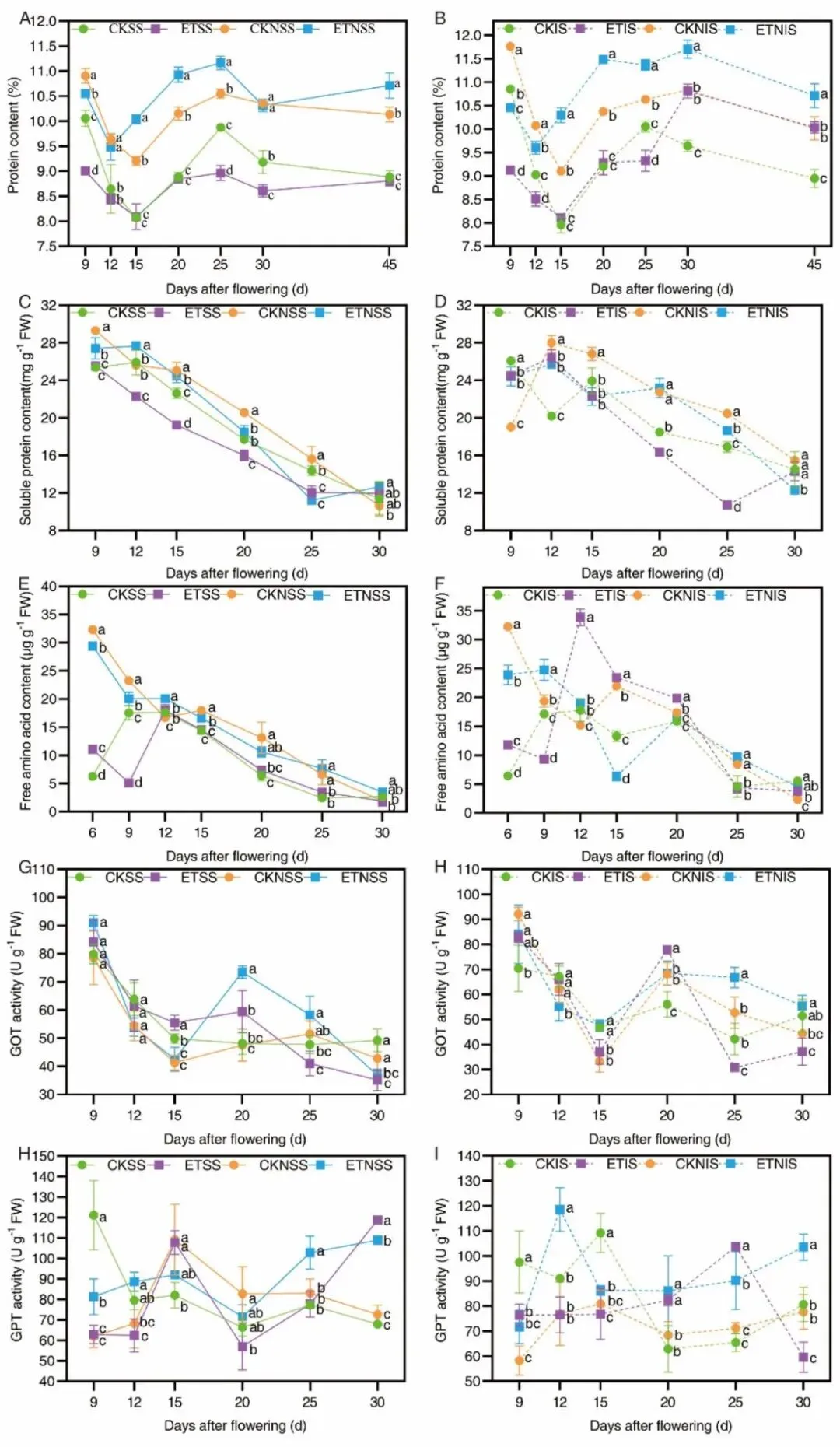
图6.强弱穗籽粒蛋白、可溶性蛋白、游离氨基酸、GOT/GPT转氨酶在全灌浆周期的动态变化。
籽粒分粒指标结果显示升温对强弱粒调控存在显著分化:强势粒粒重降2.1%、成熟期蛋白无显著变化,弱势粒粒重增4.23%、蛋白提升2.95%;籽粒可溶性蛋白、游离氨基酸随灌浆持续下降,灌浆中后期籽粒GOT、GPT转氨酶在升温与施氮诱导下活性上升,促进游离氨基酸向贮藏蛋白转化。升温环境下强势粒戊糖磷酸途径关键前体(6 -磷酸葡萄糖、6 -磷酸果糖、4 -磷酸赤藓糖)含量显著下调,NADPH合成受限、芳香族氨基酸合成受阻、内源生长素IAA下降,进而抑制强势粒灌浆充实;弱势粒则依托更高L-组氨酸、D-棉子糖、龙胆酸积累提升耐热性,规避高温代谢损伤,充分利用源叶转运氮素实现蛋白与粒重双提升。
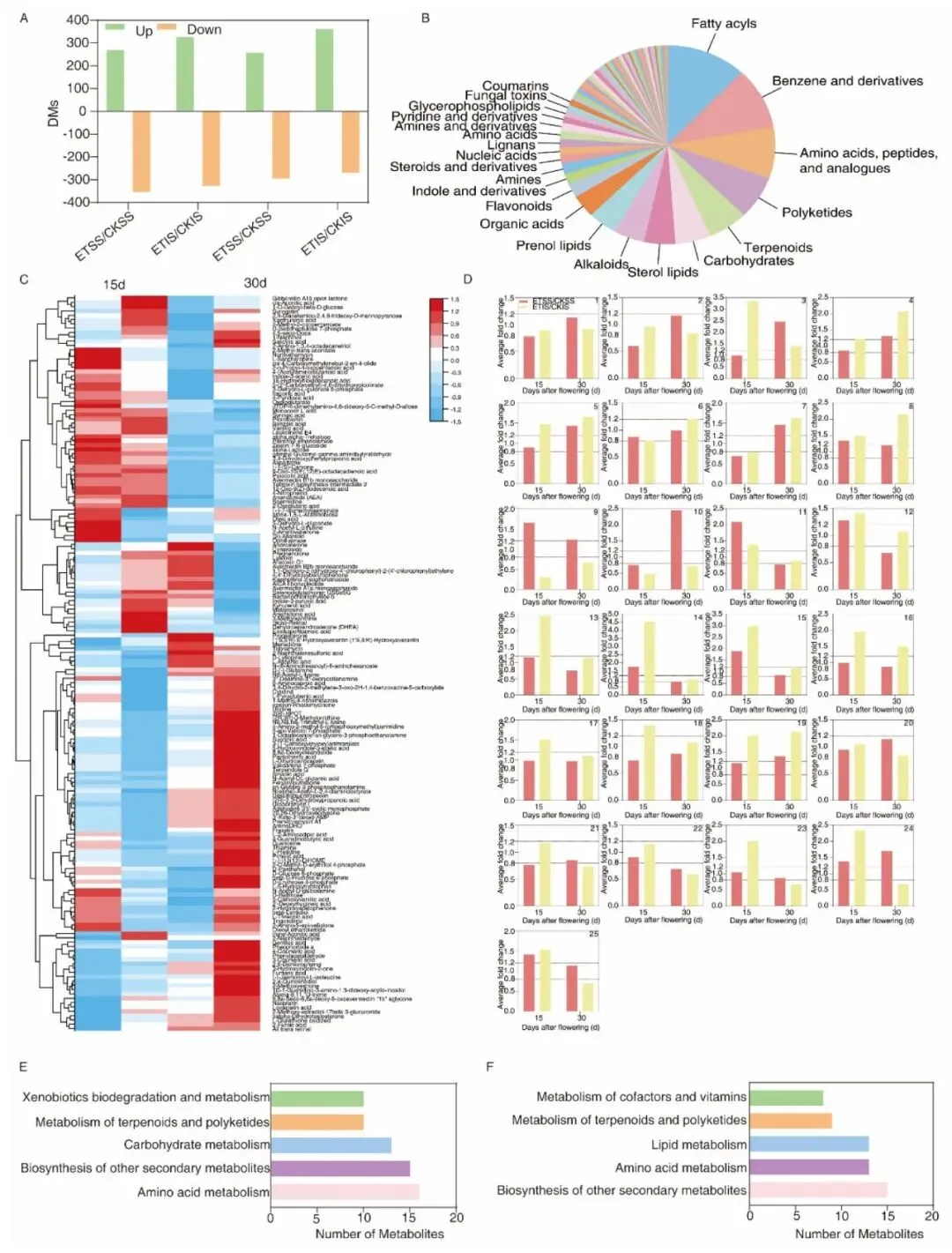
图 7.强弱粒在两个关键时期升温诱导差异代谢物数目、分类、聚类与通路富集结果。
代谢组分析选取15 DAF(籽粒蛋白低谷期)、30 DAF(灌浆近完成期)两个关键节点对旗叶、强弱粒开展LC-MS非靶向检测,共注释1773个代谢物,PCA与OPLS-DA证实温度、发育天数是代谢分异核心因素。叶片层面升温大幅重塑氨基酸、碳水、苯丙烷代谢通路,15 DAF升温上调苯丙氨酸、顺乌头酸等抗逆代谢物,30 DAF全氨基酸通路普遍受抑、叶片氨基酸大量外运耗竭;籽粒代谢上,升温显著改变色氨酸、赖氨酸、苯丙氨酸等氨基酸代谢与类黄酮、苯丙烷次生代谢,弱势粒棉子糖、龙胆酸持续高富集是其耐高温、高效合成蛋白的关键代谢特征。
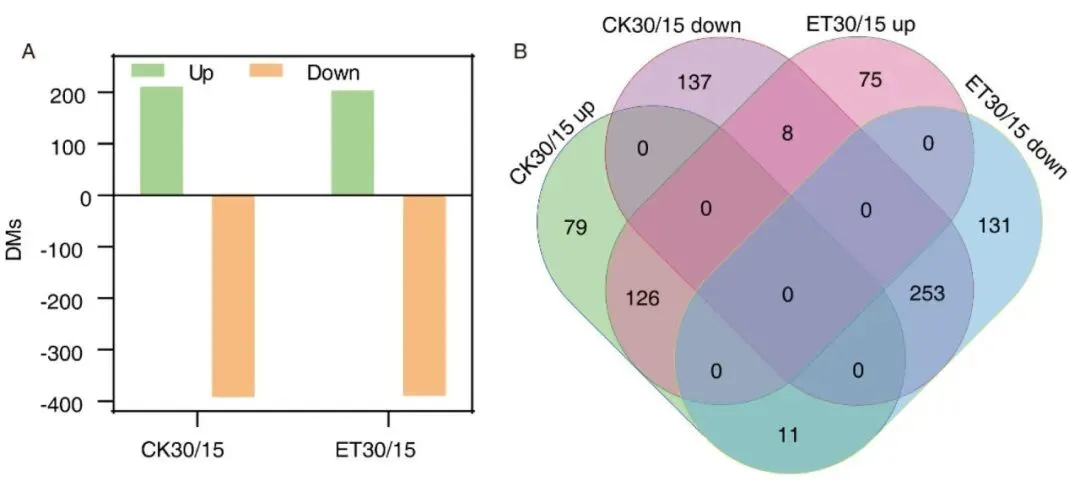
图8.常温、升温条件下叶片30 DAF/15 DAF差异代谢物数量与共有差异代谢物韦恩图。
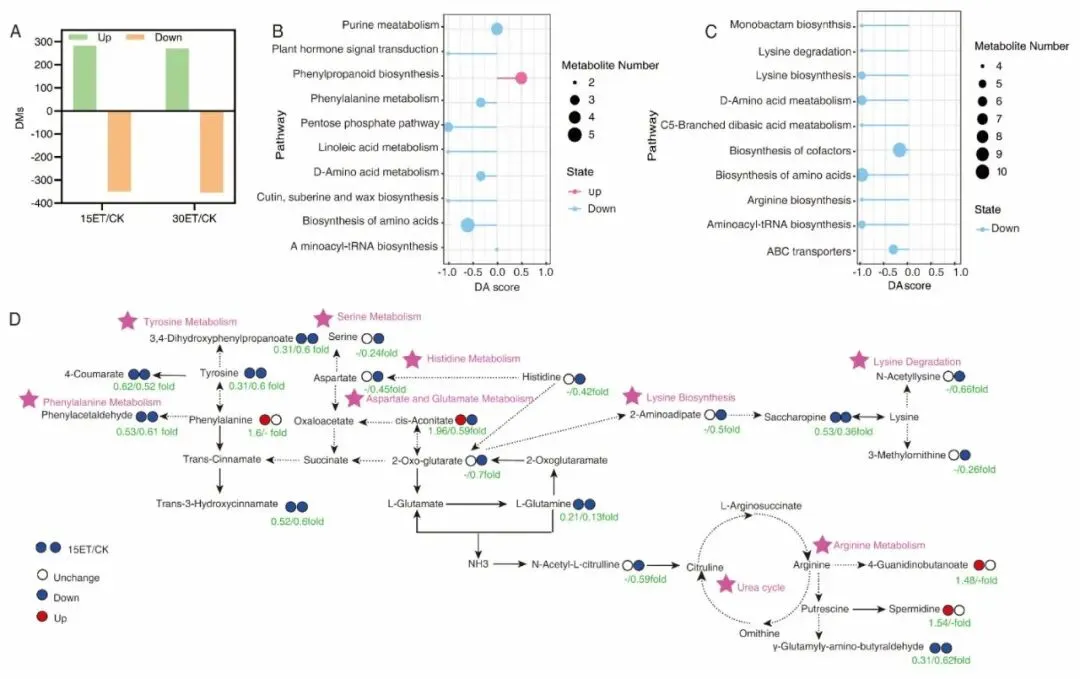
综合讨论指出,高温诱导叶片“早期氮同化增强、中后期氮外运致早衰”是强弱粒蛋白分化的核心源头机制:强势粒生理温度更高、高温胁迫更强,碳代谢受阻优先限制淀粉积累,单位籽粒蛋白占比变化有限;弱势粒穗部微环境温度偏低,高温损伤更小,源端持续输入的氨基酸在GOT/GPT催化下高效合成贮藏蛋白;外源补氮通过ROS稳态调控延缓叶片衰老、延长源氮供给周期,是同步提升两类籽粒蛋白的栽培调控路径。研究同时提出,未来可依托同位素示踪与代谢流技术量化不同器官氮转运通量,结合品种筛选与精准氮肥运筹,在稳产前提下定向提升稻米蛋白营养品质。
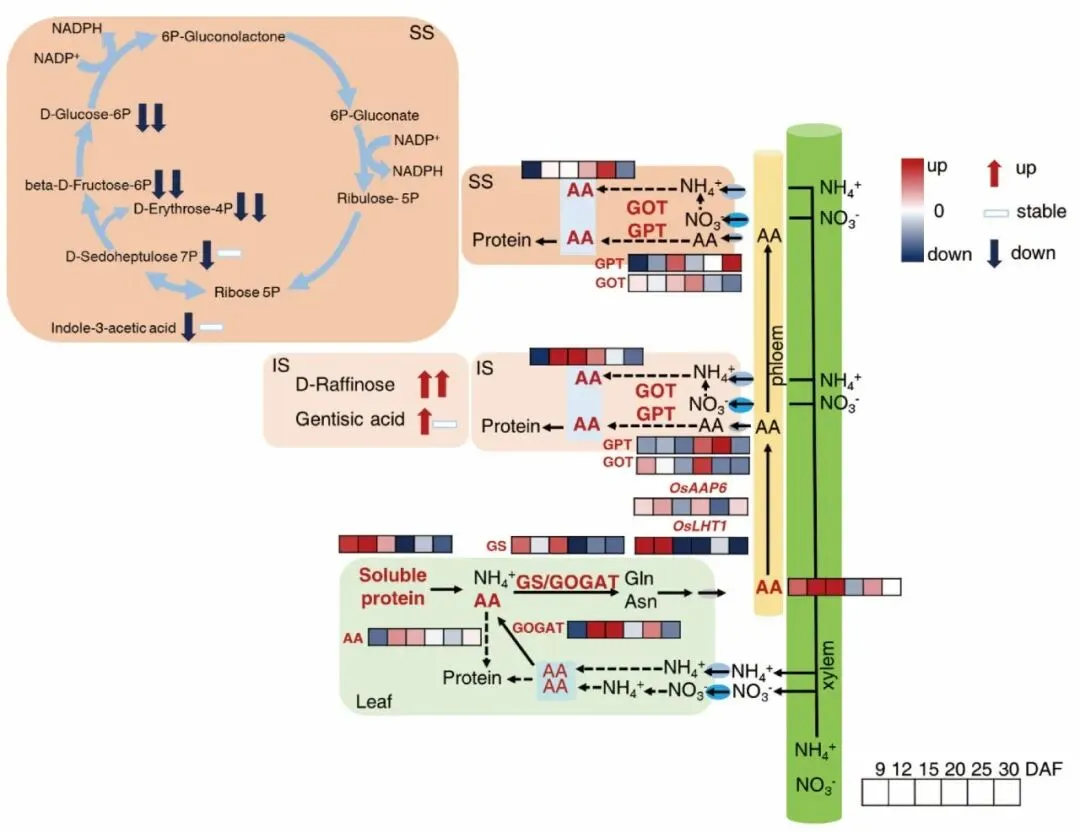
图10.全文机理总结示意图,系统梳理升温从叶片衰老 - 氮转运到强弱粒碳氮代谢分化的调控路径。
参考文献
Yufei Zhao, Chen Zhang, Wentao Shi, Ke Liu, Wei Wu, Yifan Wang, Ruiqi Li, Yuxuan Peng, Yingying Shen, Wenzhe Liu, Yanfeng Ding, Min Xi, She Tang, Elevated temperature accelerates leaf senescence and promotes leaf nitrogen transport to improve rice grain protein synthesis,Plant Physiology, 2026;, kiag310, https://doi.org/10.1093/plphys/kiag310

