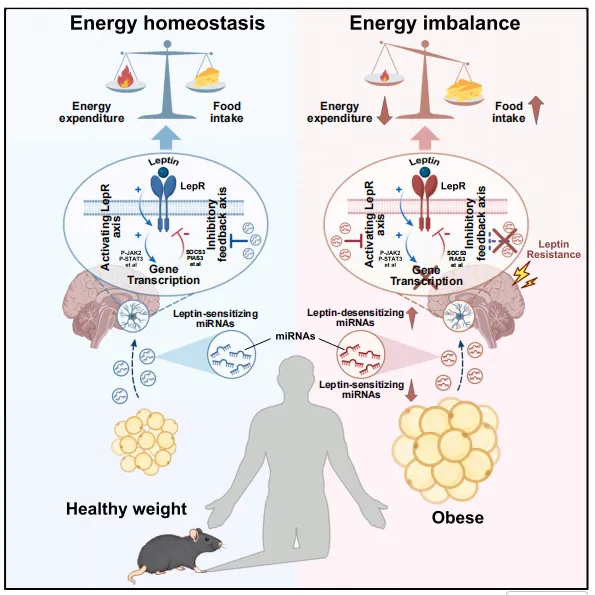
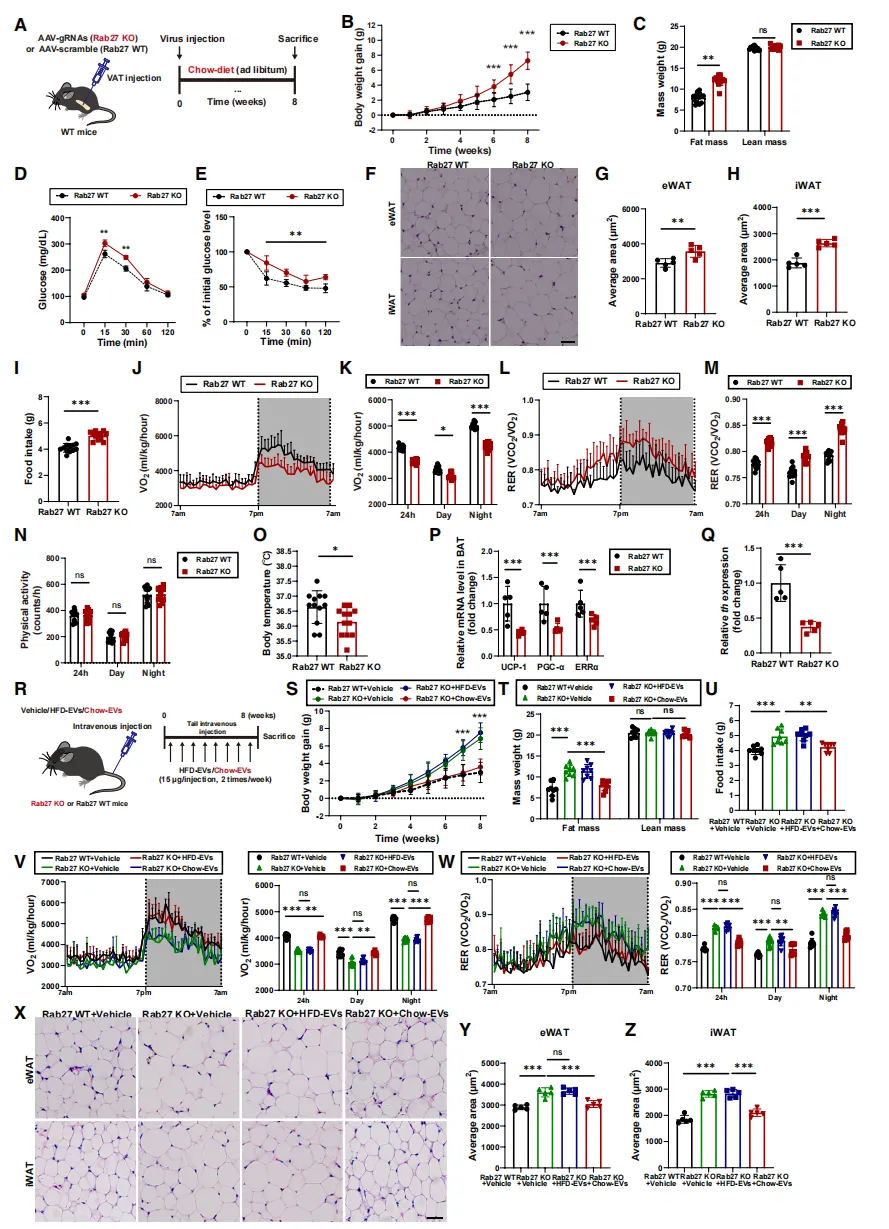
为探究脂肪组织来源细胞外囊泡(Ad-EVs)在能量代谢中的作用,研究人员构建了脂肪组织EV分泌缺陷小鼠(Rab27 KO)。
结果显示,尽管这些小鼠进食量减少,但能量消耗显著降低,体重增加更快、脂肪细胞体积明显增大【图1B-H】。
这表明Ad-EVs并非代谢过程的被动产物,而是主动参与能量平衡调控的重要信号介质。
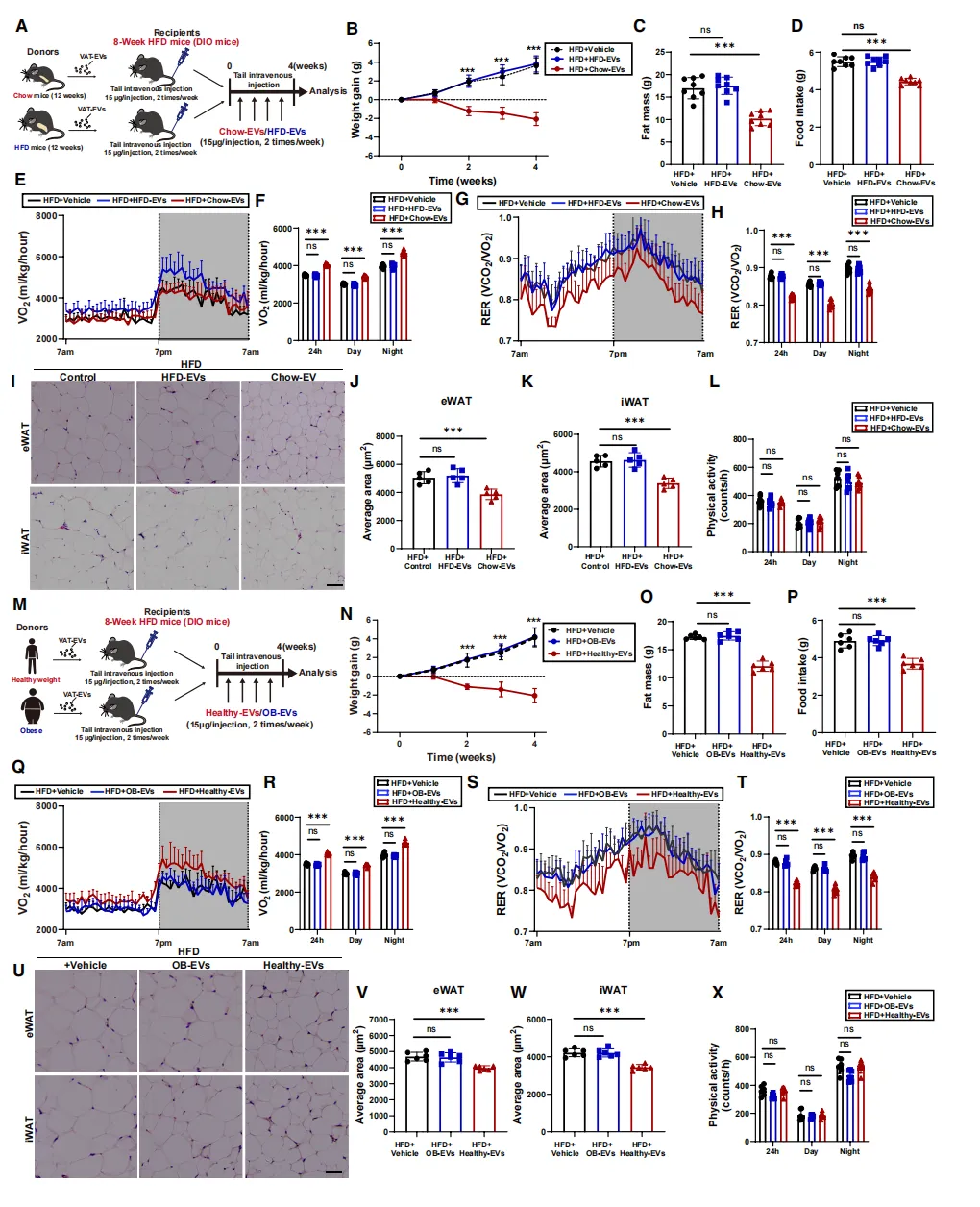
为验证这一猜想,研究人员给肥胖小鼠注射了来自瘦鼠的脂肪EV(chow-EVs)。
短短4周,效果显著:体重和脂肪量明显下降【图2B、C】,摄食减少、能耗增加【图2D-H】,脂肪细胞体积缩小【图2I-K】。
更具临床意义的是,用健康体重人群的EV治疗同样有效【图2N-X】,而肥胖来源的EV则完全无效。这说明EV的功能取决于供体的代谢状态。
关键验证:在缺乏瘦素受体的db/db肥胖鼠中,同样的EV治疗完全失效。
图3.AT-EVs对调节下丘脑瘦素-黑皮质素
信号通路至关重要
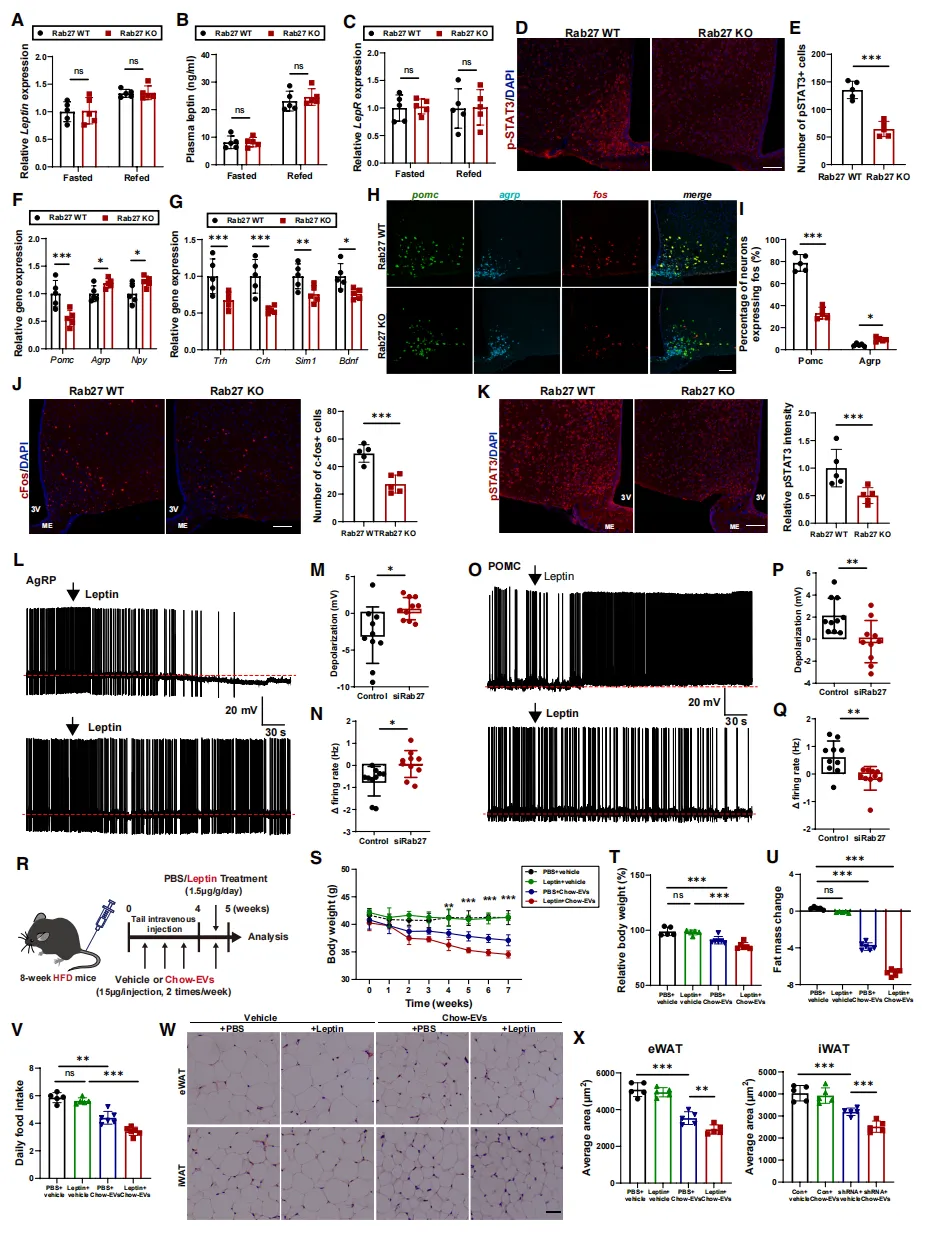
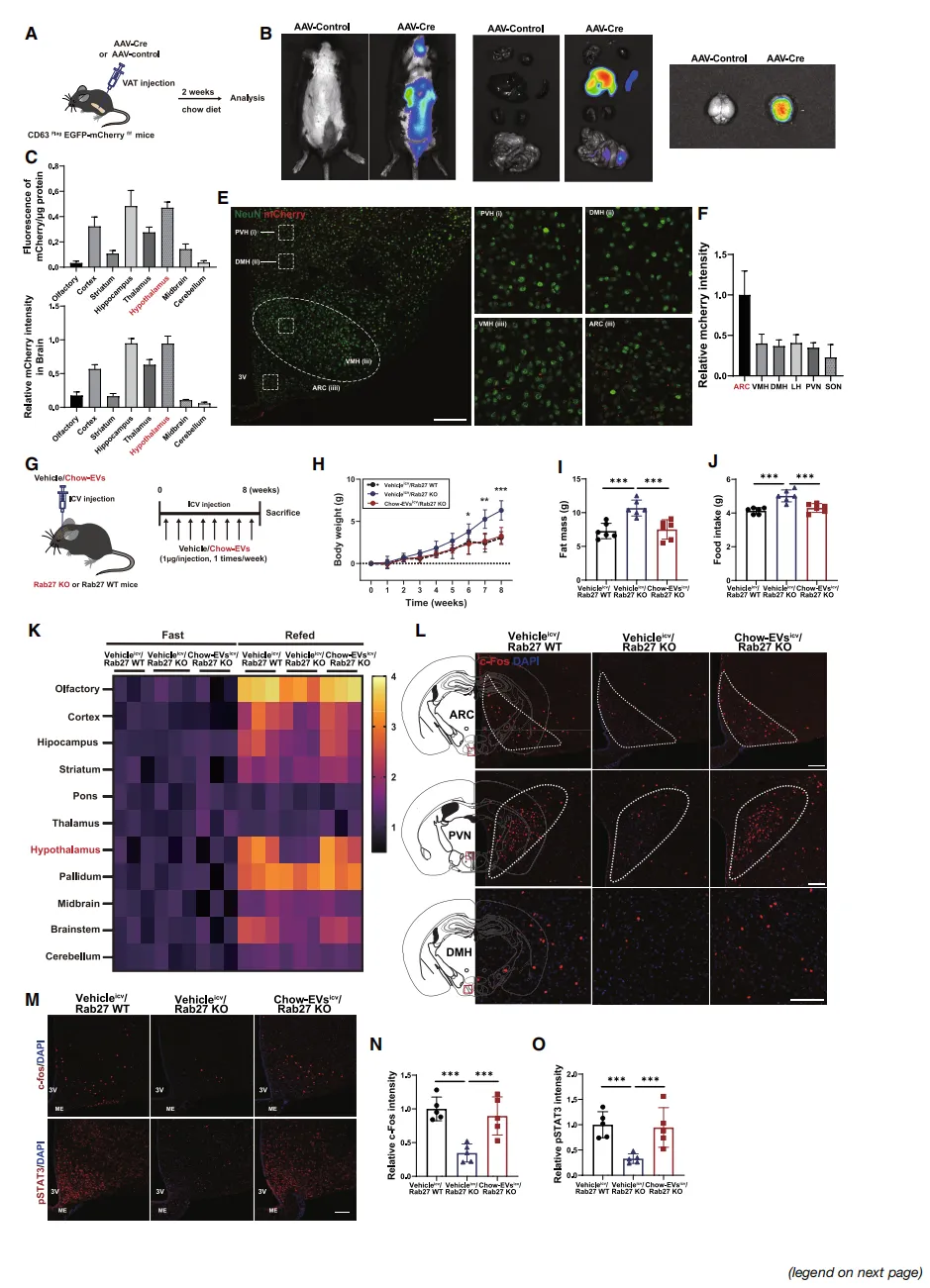
机制研究发现,脂肪EV缺乏的小鼠,大脑中感知“饱”信号的关键分子pSTAT3活性降低【图3D、E】,促进食欲的神经元(AgRP)异常活跃,而抑制食欲的神经元(POMC)反应迟钝【图3F-I】。
电生理记录直接证实:失去EV信号后,AgRP和POMC神经元对瘦素的反应均明显变差【图3L-Q】。
脂肪EV是如何与大脑对话的?荧光示踪技术给出答案:脂肪EV注入体内后,会特异性地富集在下丘脑弓状核(ARC)——调控食欲的核心区域【图4C-F】。
图4.AT-EVs直接作用于
中枢神经系统以调节能量稳态
更直接的证据:将正常EV直接注入基因缺陷小鼠的脑室,竟能“修复”全身EV缺失导致的代谢紊乱【图4H-L】。这证实脂肪EV直接作用于中枢神经系统。
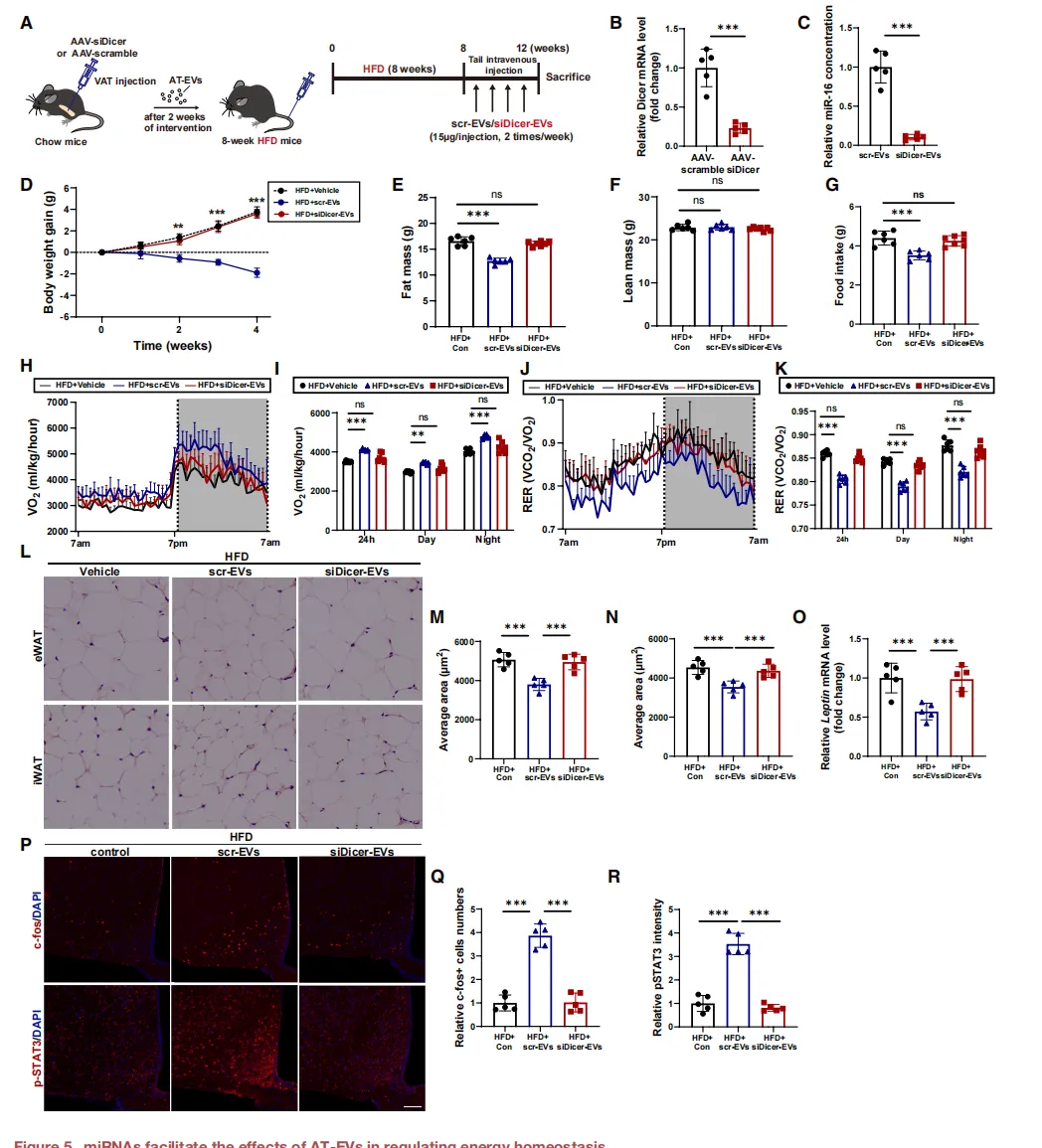
当通过敲低Dicer阻断脂肪细胞miRNA生成后,其来源EV完全丧失减肥功效【图5D-O】。
图5.miRNAs促进AT-EVs
在调节能量稳态中的作用
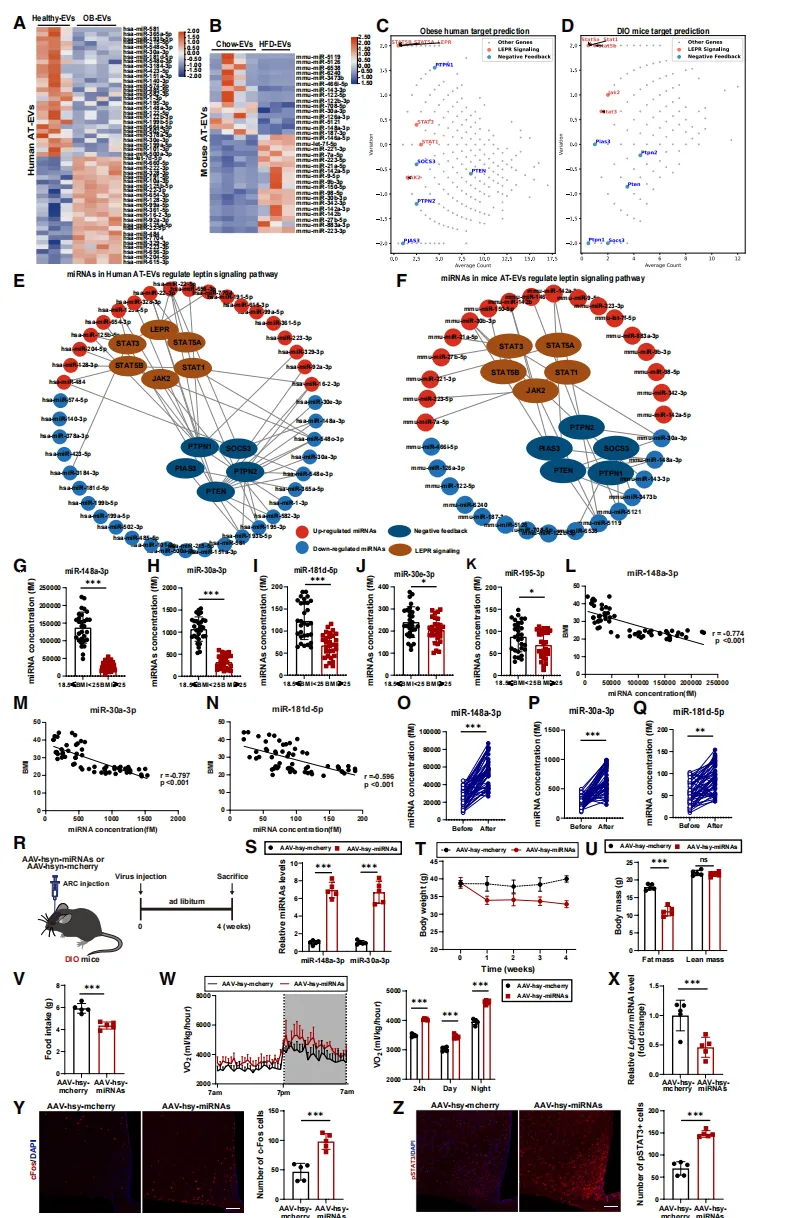
深度测序分析显示,肥胖个体EV中一群具有瘦素增敏潜能的miRNA(如miR-148a-3p、miR-30a-3p)显著下调,而可能破坏瘦素信号的miRNA则上调【图6A-F】。
图6.miRNA文件在健康-EV
功能中富集,作为瘦素增敏剂
减肥手术后,患者血清中这些“增敏”miRNA水平明显回升【图6O-Q】,提示其临床相关性。
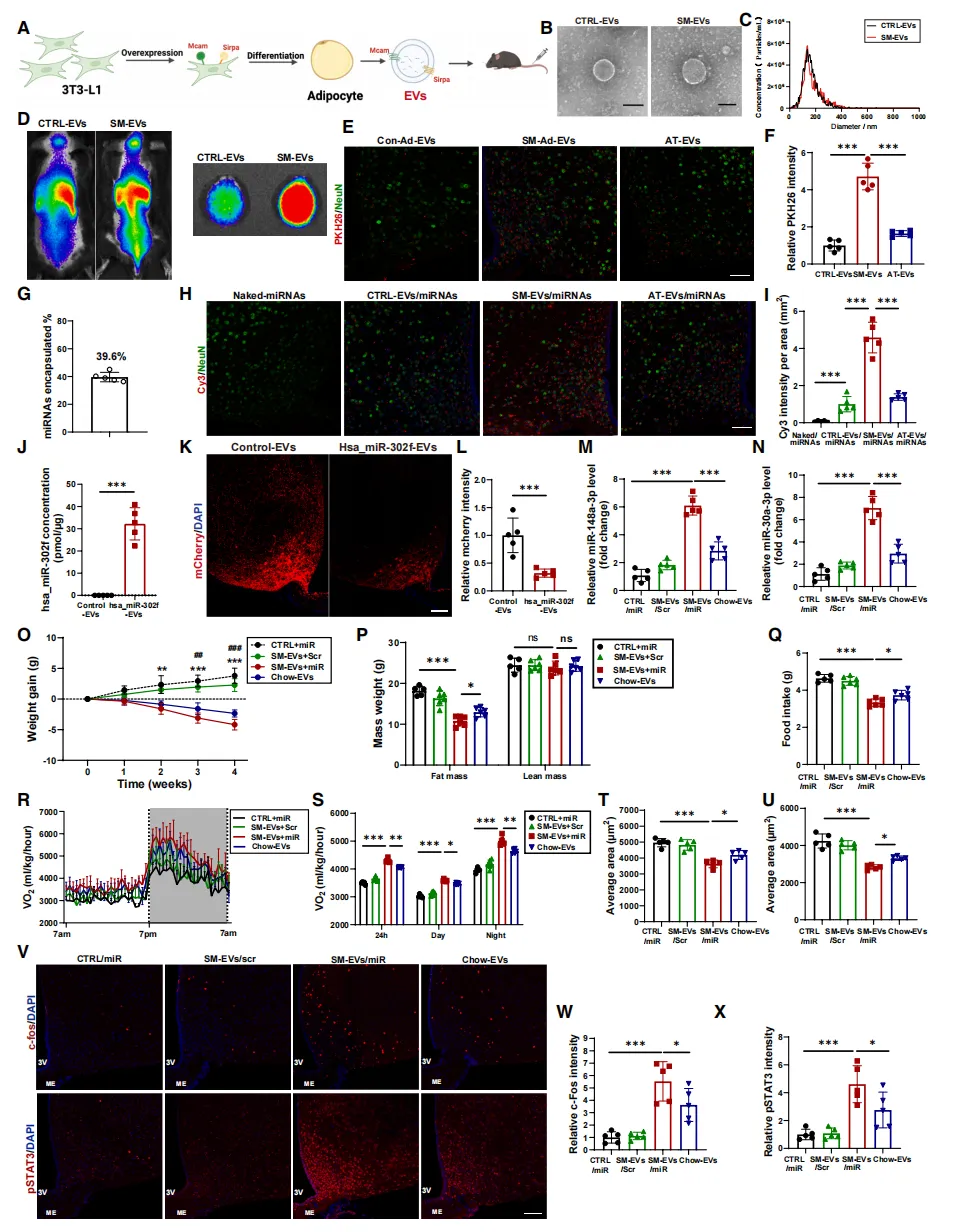
进一步研究发现,Ad-EVs表面SIRPα和MCAM蛋白是其被中枢神经系统特异性摄取的分子基础。
图7.利用工程化外泌体靶向
递送瘦素敏化miRNA缓解肥胖
基于此,研究人员构建了过表达这两种蛋白的工程化EV【图7A】,并将miR-148a-3p和miR-30a-3p装载其中。
该工程化EV能高效递送miRNA至下丘脑【图7D-I】,并在肥胖小鼠模型中恢复瘦素敏感性,显著降低体重、减少摄食、增加能耗【图7O-X】。




























 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
