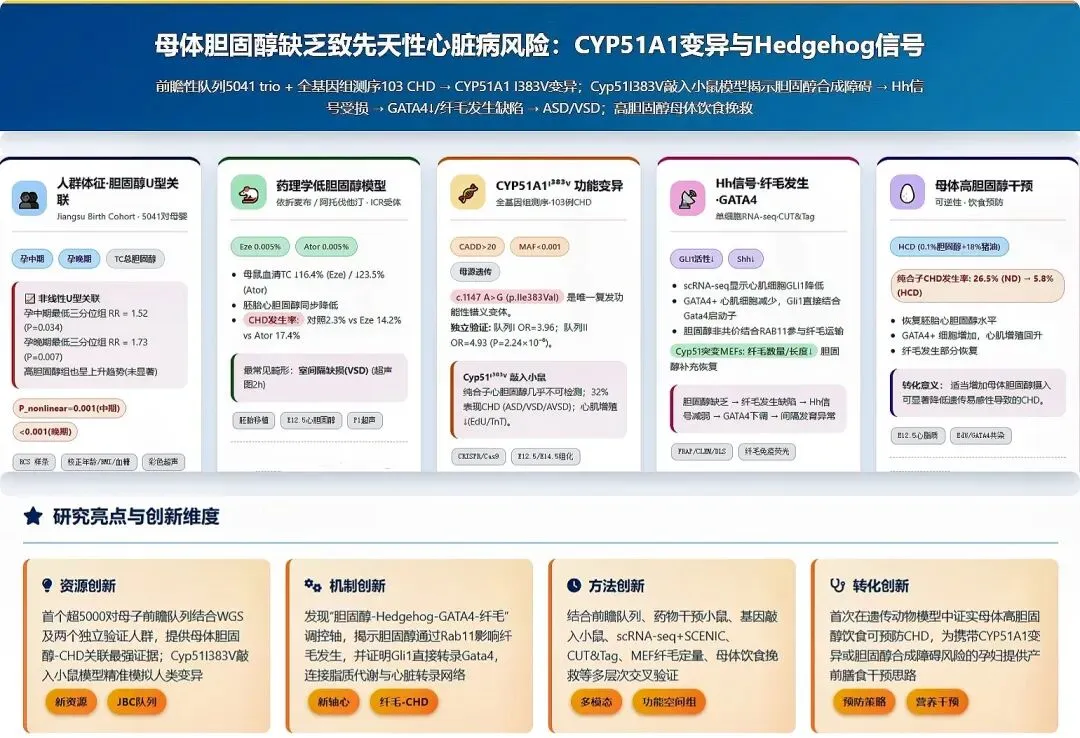
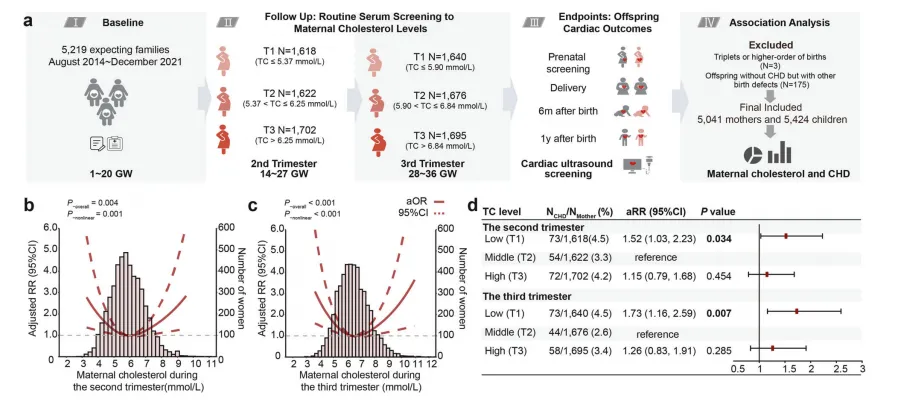
研究团队对5041对母子的出生队列数据分析显示,母体总胆固醇水平与子代先天性心脏病(CHD) 风险呈清晰的“U型”关系【图1B, C】,胆固醇过高或过低均可能增加发病风险。
其中低胆固醇群体风险尤为突出:与胆固醇水平处于中间区间(T2)的孕妇相比,妊娠中期胆固醇最低组(T1)子代CHD风险升高52%,妊娠晚期该风险进一步升至73%【图1D】。这是首次在大规模人群中证实母体胆固醇缺乏是CHD的独立风险因素。
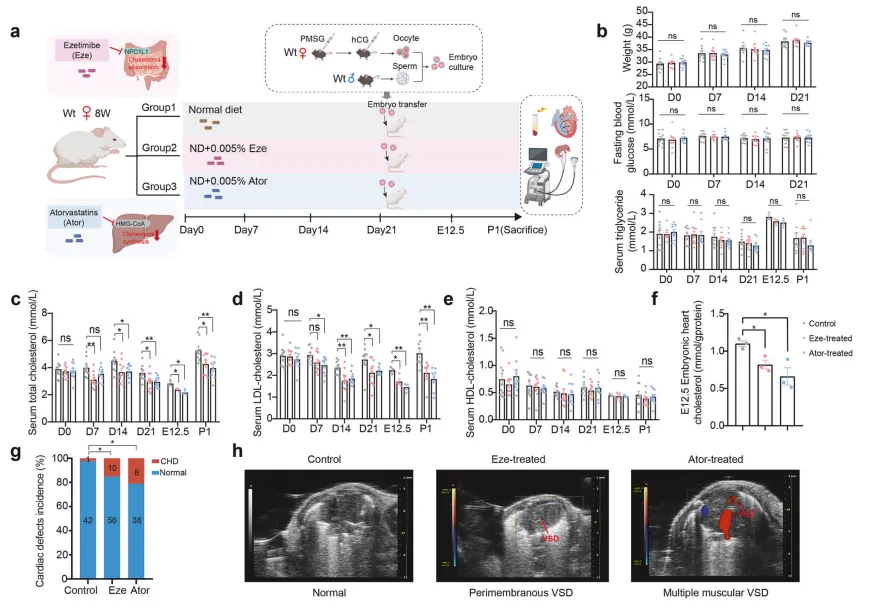
为验证人群发现的因果关系,研究人员用两种降胆固醇药物处理怀孕母鼠:依折麦布(抑制胆固醇吸收)和阿托伐他汀(抑制胆固醇合成)【图2A】。
图2.验证母体低胆固醇小鼠模型
子代冠心病风险升高的研究
结果显示,药物处理成功降低了母鼠血清及胚胎心脏的胆固醇水平【图2C-F】,且直接导致子代CHD发病率大幅上升:对照组仅为2.3%,依折麦布组升至14.2%,阿托伐他汀组高达17.4%【图2G】,证实孕期母体胆固醇短期缺乏即可显著损伤子代心脏发育。
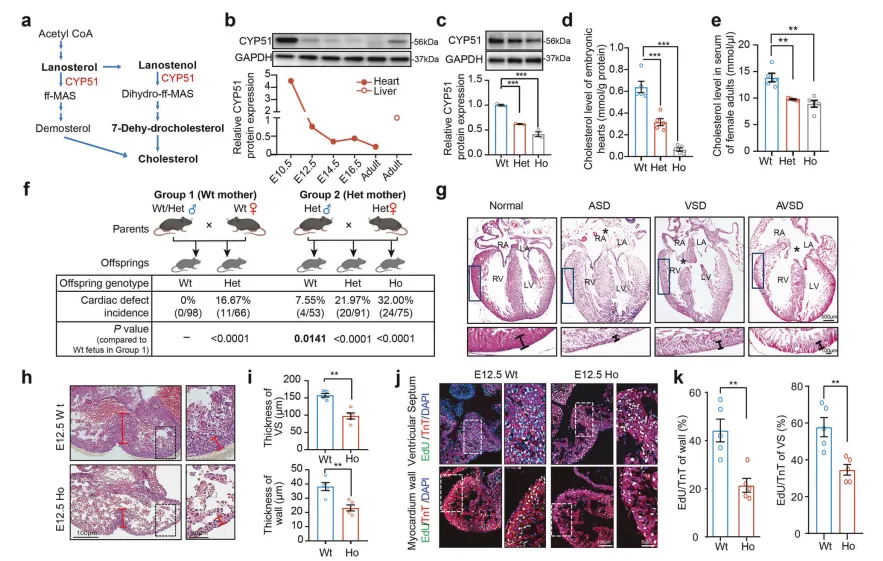
对103名CHD患儿的全基因组测序显示,胆固醇合成关键基因CYP51A1的功能性突变(I383V) 与疾病高度相关。
该突变虽罕见,但在两个独立大人群验证中均显示,携带者CHD风险为正常人的4~5倍。
图3.CYP51A1ᴵ³⁸³ⱽ基因缺陷
导致胆固醇合成受阻,引发心脏发育异常
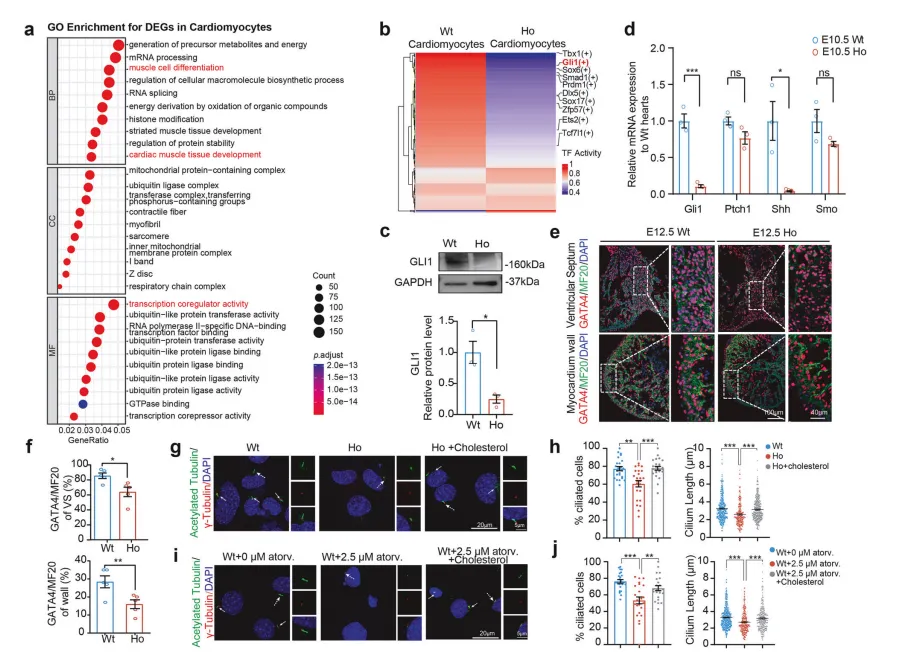
构建携带同源突变的基因敲入小鼠模型证实,该突变破坏了CYP51蛋白功能【图3C】,纯合突变小鼠胚胎心脏中胆固醇几乎检测不到【图3D】,32% 的新生纯合突变小鼠出现房间隔缺损(ASD)、室间隔缺损(VSD) 等CHD表型【图3F-H】,
核心原因是突变体心肌细胞增殖能力显著下降,导致心脏结构变薄、分隔不全【图3J, K】。
单细胞测序分析发现,突变体心肌细胞中胚胎发育核心调控通路Hedgehog的活性显著降低【图4A, B】,该通路活性高度依赖胆固醇:
图4.由CYP51A1ᴵ³⁸³ⱽ变异引起的胆固醇缺乏
导致刺猬信号通路活性受损和纤毛发生异常
直接抑制通路核心蛋白:突变体中Hedgehog通路核心效应蛋白GLI1表达量显著下降【图4C】;
破坏信号接收结构:细胞表面初级纤毛是接收Hedgehog信号的“天线”,胆固醇缺乏会导致纤毛变短、数量减少,信号接收能力大幅下降【图4G-J】。
GLI1可直接结合心脏发育关键转录因子GATA4的启动子调控其表达,最终形成“胆固醇缺乏→纤毛受损→Hedgehog信号减弱→GATA4表达下降→心脏发育缺陷”的完整致病链条【图4E, F】。
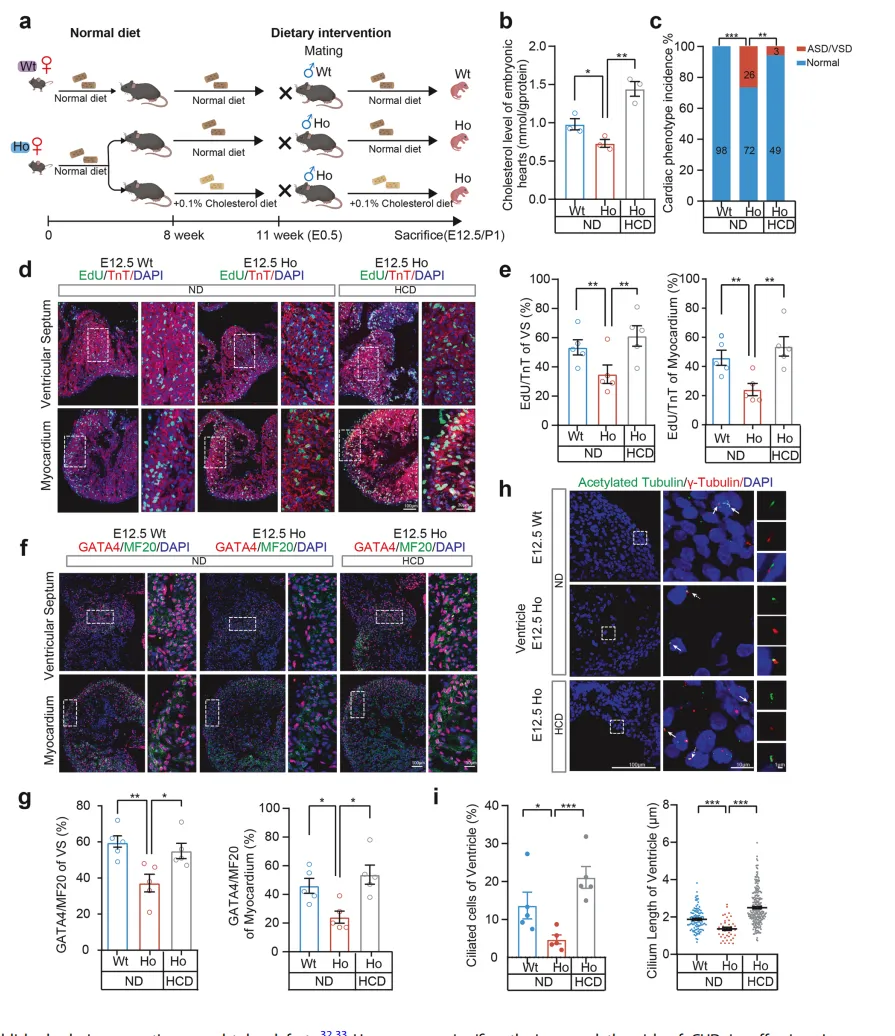
“挽救实验”显示,给携带CYP51A1ᴵ³⁸³ⱽ突变的怀孕母鼠喂养高胆固醇饮食后,纯合突变子代小鼠的CHD发病率从26.5% 大幅降至5.8%【图5C】。
图5.采用高胆固醇饮食对
携带CYP51A1ᴵ³⁸³ⱽ基因的母体进行预处理,
可降低子代先天性心脏病风险。
高胆固醇饮食不仅恢复了胚胎心脏的胆固醇水平【图5B】,还同步修复了心肌细胞增殖能力【图5D, E】、恢复了GATA4表达【图5F, G】,并挽救了纤毛发育缺陷【图5H, I】。