水稻作为全球半数以上人口的主粮,其籽粒中的储存蛋白不仅是人类膳食植物蛋白的重要来源,更是决定稻米营养品质与加工特性的关键因素。在这些储存蛋白中,谷蛋白占据了60%至80%的比例。谷蛋白的生物合成与精准分选是一个高度复杂的细胞生物学过程:其前体在内质网合成后,需经高尔基体加工,通过致密囊泡转运至蛋白储存液泡,最终被切割为成熟的酸性和碱性亚基并沉积。尽管此前学界已鉴定出多个调控这一过程的蛋白因子,但磷脂类信号分子在此过程中的具体功能与调控机制长期处于“黑箱”状态。特别是III类磷脂酰肌醇3-激酶的核心亚基VPS34,由于其纯合突变会导致生物体致死,极大地限制了科学家对其在种子发育中功能的深入解析。
近日,南京农业大学与中国农科院作物科学研究所的万建民院士、董慧教授及王益华教授团队合作,在国际植物学权威期刊《Plant Physiology》上发表了突破性研究成果。该研究利用筛选到的谷蛋白前体积累弱突变体gpa14,结合图位克隆、脂质组学及细胞生物学等多维度技术手段,首次阐明了磷脂酰肌醇3-磷酸(PI3P)在谷蛋白转运中的核心作用,揭示了OsVPS34介导的PI3P合成如何通过招募调控因子GPA5/Rab5a,促进致密囊泡与蛋白储存液泡的融合,从而保障谷蛋白正确沉积的分子机制。
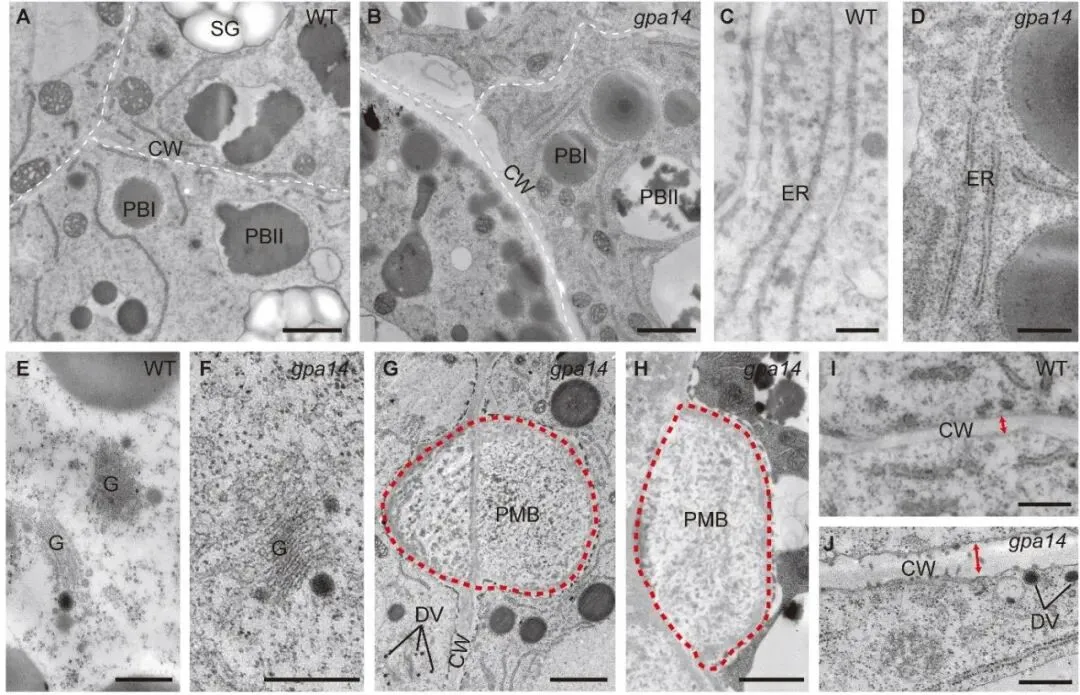
研究团队首先从粳稻品种宁粳4号的突变体库中筛选获得了gpa14突变体。该突变体表现出显著的粉质皱缩表型,扫描电镜观察显示其胚乳淀粉颗粒呈球形且排列松散。生化分析表明,突变体中57 kDa的谷蛋白前体大量积累,而成熟的谷蛋白亚基及α-球蛋白含量显著降低。值得注意的是,内质网分子伴侣的含量在突变体与野生型中无显著差异,这排除了内质网输出缺陷的可能性,暗示问题出在高尔基体后的转运环节。细胞学观察进一步证实了这一推断:在野生型中,谷蛋白与α-球蛋白共同定位于蛋白储存液泡II中;而在gpa14突变体中,部分蛋白被错误分选至胞外形成副壁体结构,且蛋白储存液泡II的体积显著缩小。透射电镜结果显示,突变体中高尔基体出芽形成的致密囊泡无法正常转运,而是堆积在细胞膜附近并被分泌到胞外。
通过图位克隆技术,研究人员将目标基因锁定在5号染色体的特定区间,并发现Os05g0180600基因存在单碱基插入导致的移码突变。该基因编码III类磷脂酰肌醇3-激酶的催化亚基OsVPS34。遗传互补实验证实,在突变体中过表达全长OsVPS34可完全恢复其表型,确证了该基因的功能。进化分析与表达模式研究显示,OsVPS34在植物界高度保守,且在水稻发育胚乳中持续高表达,与谷蛋白的合成时期高度吻合。
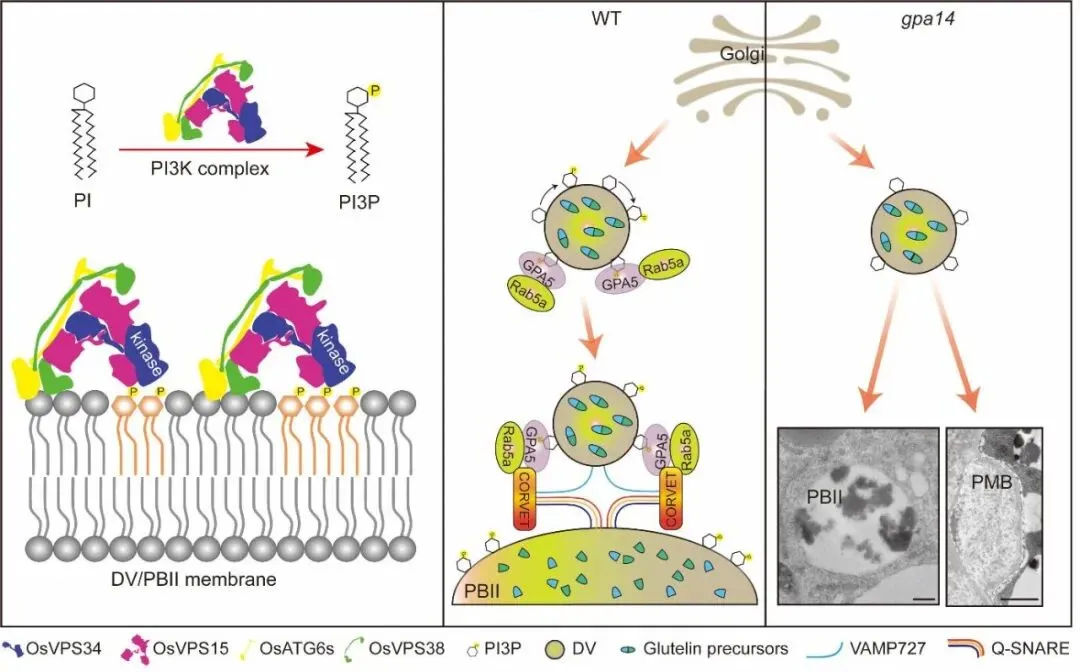
为了解析OsVPS34的作用机理,研究团队通过免疫共沉淀联合质谱技术,鉴定出OsVPS34与OsVPS15、OsATG6b、OsVPS38等蛋白互作,证明水稻中存在调控自噬和囊泡转运的两类PI3K复合体。实验证实,OsVPS34与OsATG6b形成的复合体共定位于反式高尔基体网络和前液泡区室,这正是谷蛋白转运的关键细胞器。作为PI3K复合体的核心功能,PI3P的合成在突变体中受到严重阻碍,其含量较野生型降低了60%以上。利用特异性报告系统的共定位分析进一步证实,PI3P主要富集于反式高尔基体网络、前液泡区室以及致密囊泡中,直接参与了谷蛋白的转运过程。
深入的研究揭示了PI3P的下游调控机制。GPA5作为Rab5a的效应子,其N端PX结构域负责结合PI3P。细胞组分分离实验发现,在野生型中GPA5主要定位于膜组分,而在gpa14突变体中,膜结合型GPA5含量显著降低。使用PI3K抑制剂处理或构建诱导型RNAi株系均证实,缺乏PI3P会导致Rab5a和GPA5无法正确锚定在膜系统上,从而阻断了致密囊泡与蛋白储存液泡的融合。
基于上述发现,研究团队提出了完整的分子工作模型:在水稻胚乳中,OsVPS34作为核心催化亚基,在特定细胞器膜上催化合成PI3P;PI3P作为关键的膜信号分子,招募效应子GPA5和小G蛋白Rab5a至膜表面;随后,GPA5通过与栓系复合体及SNARE融合复合体互作,介导囊泡融合,完成谷蛋白的转运与沉积。若OsVPS34功能缺失,这一链条断裂,导致谷蛋白被错误分泌。
这项研究不仅克服了核心亚基致死的研究瓶颈,首次直接证明了磷脂信号分子PI3P在水稻谷蛋白转运中的核心功能,完善了种子储存蛋白分选的调控网络,还为稻米品质的分子改良提供了直接靶点。通过编辑调控该通路,未来有望定向优化谷蛋白含量,培育出高营养或适合特定人群(如肾病患者)食用的功能性稻米。同时,这一保守的调控模块也为小麦、玉米等其他禾本科作物的品质改良提供了新的理论方向。