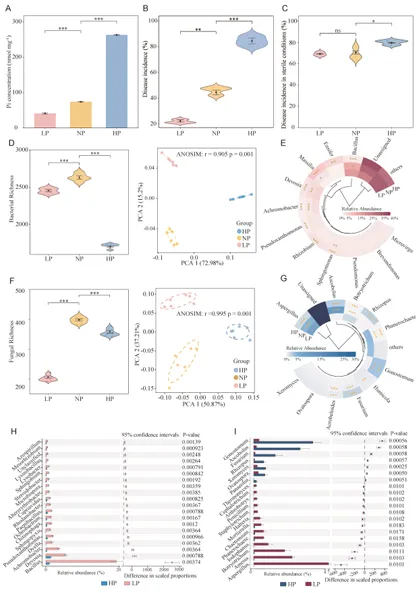
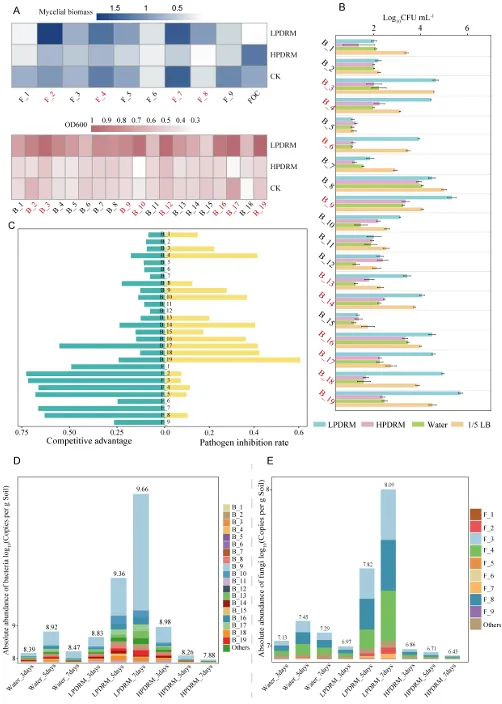
Microbiome | 南京农业大学:磷有效性通过调控根际代谢物-微生物群落互作影响黄瓜对枯萎病的易感性
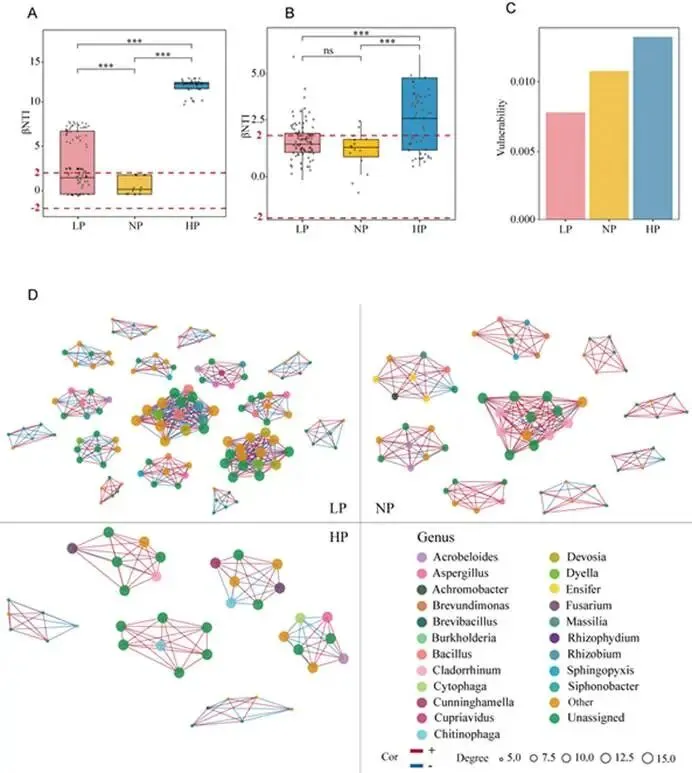
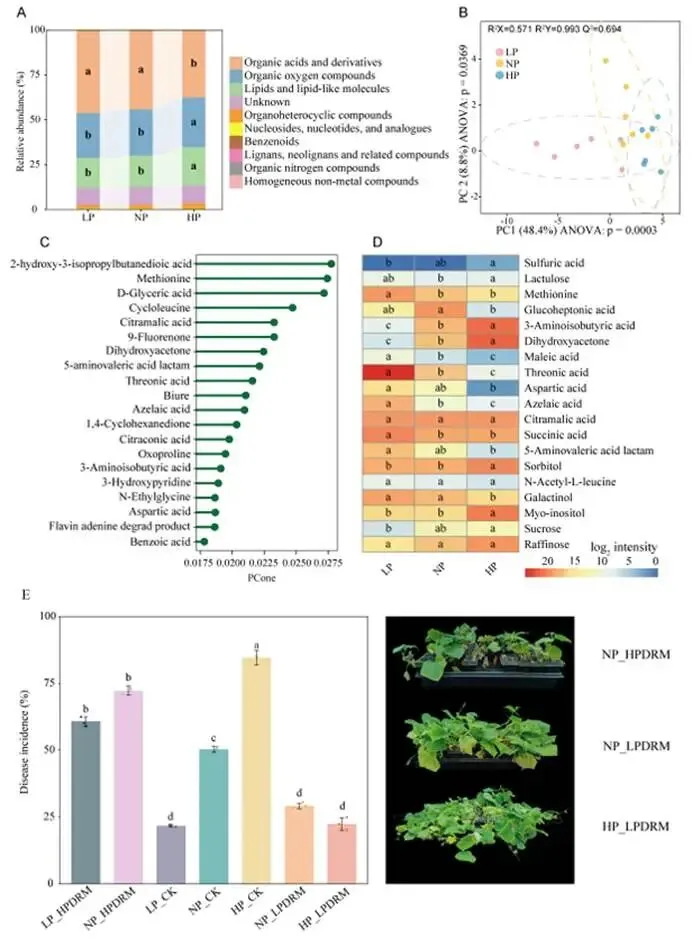
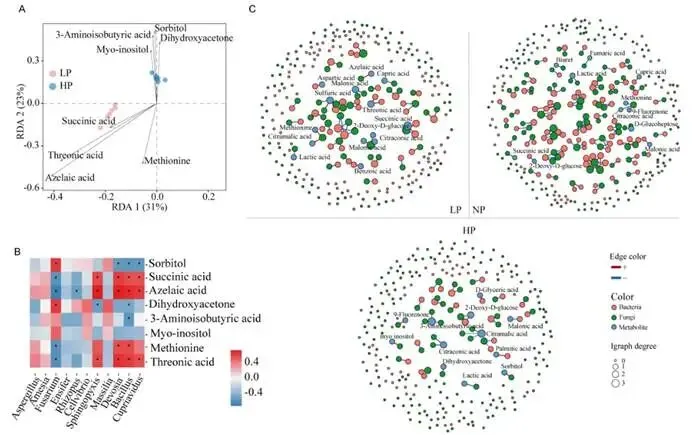
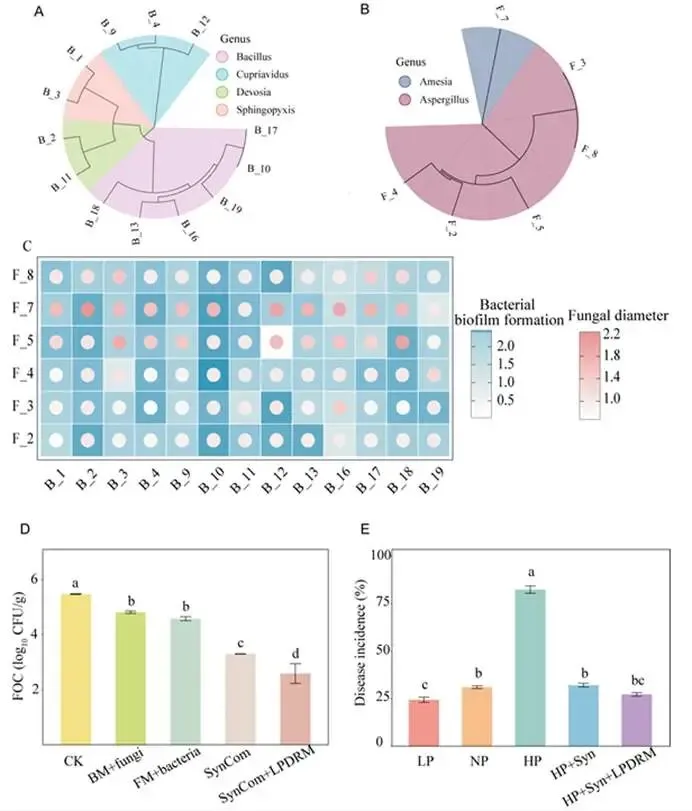
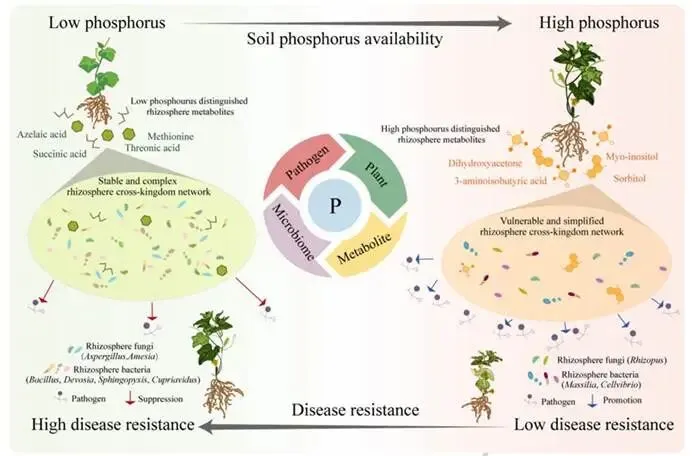
背景:农业集约化导致农田磷肥过量施用,然而土壤磷富集与由根际微生物群落介导的土传病害易感性之间的因果关系尚不明确。本研究旨在解析磷有效性如何通过重构根际代谢物-微生物群落互作来调控黄瓜对枯萎病的易感性。结果:本研究证实,适度低磷条件可增强黄瓜对枯萎病的抗性。扩增子测序分析表明,低磷环境促进微生物群落的随机组装,富集兼具解磷与抗病双重功能的有益菌属(细菌属包括芽孢杆菌属 Bacillus、德沃斯氏菌属 Devosia、鞘氨醇单胞菌属 Sphingopyxis、贪铜菌属 Cupriavidus;真菌属包括曲霉属 Aspergillus 和Amesia属),并增强跨界网络鲁棒性。相比之下,高磷条件引发确定性组装,其病害发生率较低磷处理增加62%。代谢组学分析发现,低磷条件下特征性根际代谢物(琥珀酸、壬二酸、苏糖酸和甲硫氨酸)可招募有益类群,而高磷特征代谢物则促进病原菌生长。为验证微生物-代谢物互作的功能机制,我们构建了由低磷微生物群标志性类群组成的合成群落,当其与低磷代谢物联用时,可使病原菌丰度降低85%,并在高磷条件下实现病害抑制。结论:本研究揭示了一种磷驱动的植物-代谢物-微生物群落互作协调根际免疫的机制。这些发现为具有历史磷累积的农业系统提供了可持续病害管理的新见解和潜在策略。篇名: Phosphorus availability drives rhizosphere metabolite-microbial community interactions to modulate cucumber susceptibility to Fusarium wiltDOI: 10.1186/s40168-026-02381-1该研究以黄瓜品种为“长春密刺”为材料,通过设置低、中、高三个磷浓度(500 μM、1 mM、2 mM)进行温室盆栽试验,探究磷供应对黄瓜枯萎病发生的影响,并开展无菌条件下验证微生物介导效应。采用16S rRNA及ITS扩增子高通量测序分析根际微生物群落,利用GC-TOF-MS检测根际代谢物,同时分离鉴定关键微生物菌株,构建合成菌群以评估其抑病功能。为探究磷营养对黄瓜枯萎病的影响,本研究以黄瓜(品种:长春密刺)为材料,在三种不同磷供应水平下接种尖孢镰刀菌黄瓜专化型(Fusarium oxysporum f. sp. cucumerinum, FOC)进行侵染实验。结果表明,高磷(HP)处理植株的病害易感性显著增强,发病率达84.5%,较正常磷(NP)条件高40%;而低磷(LP)处理的黄瓜发病率仅为22.2%,较正常磷处理降低22.3%(图1A、B;附图1)。为验证根际微生物群落是否参与磷水平与黄瓜枯萎病发生的关联,我们在无菌培养的黄瓜根际接种FOC病原菌。结果显示,LP与NP条件下的发病率无显著差异(分别为69.3%和70.6%;p > 0.05),而HP植株的发病率显著升高至79.7%(p < 0.05)(图1C),表明低磷条件下根际微生物群落对枯萎病具有抑制作用。进一步分析三种磷水平下黄瓜根际细菌群落发现:与NP相比,LP与HP处理的基于丰富度的α多样性均显著降低。主成分分析(PCA)显示处理间群落结构明显分离(累计解释方差88.18%),ANOSIM验证磷水平对β多样性具有强效影响(R = 0.905,p = 0.001;图1D)。变形菌门在所有根际样本中均占优势。相较于NP根际中拟杆菌门(11.57%)的显著富集,HP与LP处理中该门相对丰度均大幅降低。与LP组相比,HP组放线菌门(7.92%)相对丰度显著升高,而拟杆菌门(6.95%)与厚壁菌门(2.64%)显著降低(附图2)。根际细菌属水平热图显示,随着磷水平递减,芽孢杆菌属(Bacillus,18.86%)显著富集;反之,马赛菌属(Massilia,0.41%)相对丰度随磷水平升高而增加(图1E)。对根际真菌群落的分析表明,LP处理显著降低了基于丰富度的α多样性。PCA显示前两主成分共解释88.08%的变异(PC1:50.87%;PC2:37.21%),磷水平显著改变真菌群落结构(ANOSIM R = 0.995,p = 0.001;图1F)。子囊菌门与担子菌门在所有处理中均占优势。相较于HP与NP处理,LP条件下子囊菌门(16.03%)、担子菌门(4.00%)和球囊菌门(2.32%)相对丰度显著更高(附图3)。属水平分析显示,随着磷水平降低,优势根际真菌呈现分化响应:曲霉属(Aspergillus,7.04%)丰度显著增加,而根霉属(Rhizopus,< 0.01%)、镰刀菌属(Fusarium,0.08%)及殖口虫属(Gonostomum,0.29%)丰度显著下降(图1G)为鉴定不同磷水平下的优势微生物,采用TMM法进行差异丰度分析。微生物群落分析揭示:LP条件下细菌属(如芽孢杆菌属、德沃斯氏菌属)及真菌属(如曲霉属、Amesia属)显著富集;HP组则富集殖口虫属、粪盘菌属(Ascobolus)、镰刀菌属和根霉属(图1H、1I)。通过随机森林分析筛选与磷营养水平显著相关的微生物特征,最终确定40个最具判别力的微生物特征。基于这些特征,真菌与细菌群落均实现完全判别(准确率100%)。完整的判别微生物特征列表(含重要性排名、分类信息及组间差异丰度)详见附表3-4及附图4-5。图1. 磷依赖性病害发生模式与根际微生物群落结构。A:黄瓜地上部磷含量(n=6个生物学重复;均值±标准误)。B:不同磷水平下黄瓜枯萎病发病率(n=6个生物学重复;均值±标准误)。C:无菌对照条件下不同磷水平处理的黄瓜发病率(n=6个生物学重复;均值±标准误)。D:不同磷水平黄瓜根际细菌群落的α多样性(丰富度指数)与β多样性(PCA排序图)。箱线图周边散点为原始数据,箱体内横线为中位数,箱体上下缘分别代表第75和第25百分位数。PCA分析显示前两主成分累计解释88.18%变异(PC1:72.98%,PC2:15.20%),ANOSIM验证磷水平显著影响群落结构(R=0.905,p=0.001)。E:细菌属水平相对丰度热图。颜色深浅反映相对丰度,差异丰度菌属通过负二项广义线性模型鉴定(FDR<0.05)。星号表示模型框架内与低磷组(LP)的配对比较显著性:*p<0.05,**p<0.01,***p<0.001。F:不同磷水平黄瓜根际真菌群落的α多样性(丰富度指数)与β多样性(PCA排序图)。箱体内横线为中位数,箱体上下缘分别代表第75和第25百分位数。PCA分析显示前两主成分累计解释88.08%变异(PC1:50.87%,PC2:37.21%),ANOSIM验证磷水平显著影响群落结构(R=0.995,p=0.001)。G:不同处理真菌属水平相对丰度热图。颜色深浅反映相对丰度,差异丰度菌属通过负二项广义线性模型鉴定(FDR<0.05)。星号表示模型框架内与低磷组(LP)的配对比较显著性:*p<0.05,**p<0.01,***p<0.001。H-I:STAMP分析鉴定的差异丰度细菌属(H)和真菌属(I)(FDR校正p<0.05)。柱状图显示LP与HP组的log2倍变化值。(2)不同磷水平下根际微生物群落组装与网络驱动因子的鉴定通过β-最近分类单元指数(βNTI)分析磷梯度下的根际微生物群落发现:高磷(HP)条件下细菌与真菌群落均呈现确定性组装(βNTI > 2);而低磷(LP)与正常磷(NP)处理则以随机过程主导,|βNTI| < 2表明中性过程占优(图2A-2B)。跨界网络分析显示,HP与LP处理下黄瓜根际真菌-细菌互作网络的边数均增加,但关键差异在于:LP网络中存在显著更多的真菌-细菌负相关边,而HP网络以正相关边为主(附表5)。脆弱性评估进一步表明,HP处理的根际网络结构脆弱性显著高于NP与LP条件,其中LP跨界网络稳定性最高(图2C)。对跨界微生物网络的模块化分析揭示,随着磷有效性降低,黄瓜根际网络的模块复杂性逐渐增加。LP处理主要模块中的关键类群包括细菌属芽孢杆菌属(Bacillus)、德沃斯氏菌属(Devosia)、鞘氨醇单胞菌属(Sphingopyxis)和贪铜菌属(Cupriavidus),以及真菌属曲霉属(Aspergillus);而HP网络则表现为马赛菌属(Massilia)、根霉属(Rhizopus)和镰刀菌属(Fusarium)具有更高的拓扑中心性(图2D)。通过整合差异丰度分析、机器学习定义的判别贡献度(最具区分力的类群)及网络模块重要性评估,本研究将细菌属芽孢杆菌属、德沃斯氏菌属、鞘氨醇单胞菌属、贪铜菌属,以及真菌属曲霉属和Amesia属确定为LP组的标志性微生物类群,这些类群可能在磷限制条件下参与黄瓜枯萎病抗性调控;反之,细菌属马赛菌属和纤维弧菌属(Cellvibrio),以及真菌属根霉属被鉴定为HP组的标志性类群(附表6),这些微生物可能促进高磷条件下枯萎病严重度的增加。图2. 磷梯度下根际微生物群落的组装机制与网络分析。A-B:细菌(A)与真菌(B)群落的β-最近分类单元指数(βNTI)。|βNTI| > 2表示确定性组装(高磷处理组HP占主导);|βNTI| < 2表示随机性组装(低磷处理组LP/正常磷处理组NP占主导)。C:跨界(真菌-细菌)关联网络的脆弱性指数。数值越低表示稳定性越高。D:跨界共现网络中关键类群的拓扑学角色。节点大小反映中介中心度;颜色表示属级分类归属。LP:低磷处理;NP:正常磷处理;HP:高磷处理。为深入解析磷营养、植物与根际微生物间的互作关系,本研究采用GC-TOF-MS对根际代谢物进行分析。所有样本共鉴定出336种化合物,包括:2种苯环类、1种同质非金属化合物、1种木脂素或新木脂素类、21种脂质及类脂分子、4种核苷酸及其类似物、33种有机酸及其衍生物、1种有机氮化合物、34种有机氧化合物、6种有机杂环化合物、36种其他化合物及196种未鉴定物质。其中,高磷(HP)根际中有机酸及其衍生物相对丰度显著降低,而有机氧化合物与脂质类分子显著增加(图3A)。偏最小二乘判别分析(PLS-DA)显示三组实验处理(P1mM、P2mM、P500μM)的代谢谱存在明显分离。得分图(图3B)表明第一主成分(PC1)解释了代谢组数据48.4%的变异,第二主成分(PC2)解释8.8%,前两主成分累计解释率达57.1%。ANOVA验证组间分离在PC1(p = 0.0003)与PC2(p = 0.0369)上均具统计学显著性。载荷图分析筛选出对PC1贡献最大的前20种代谢物(图3C)。随机森林分析进一步鉴定出区分三组的前40种潜在关键代谢物(附图6),如半乳糖酸、9-芴酮、2-脱氧-D-葡萄糖和壬二酸等因其高判别贡献度被识别,模型准确率达72.2%(附图6)。非参数检验揭示HP与LP组间33种代谢物存在显著差异(p < 0.05;附表7),图3D展示组间差异最显著的前20种代谢物。综合载荷图、随机森林及非参数检验结果,本研究将琥珀酸、壬二酸、苏糖酸和甲硫氨酸定义为低磷特征根际代谢物(LPDRM),而肌醇、二羟基丙酮、山梨醇和3-氨基异丁酸定义为高磷特征根际代谢物(HPDRM)以供后续研究。为验证所选特征代谢物是否与黄瓜在不同磷水平下对枯萎病的差异抗性相关,我们设计了双因子盆栽实验(磷水平×代谢物添加)。结果表明:添加HPDRM的黄瓜植株枯萎病发病率始终维持高位(接近HP处理水平),且不受环境磷水平影响;而LPDRM添加则能显著抑制所有磷水平下的病害发展(图3F)。图3. 不同磷水平下根际代谢物的谱分析与功能验证。A:各处理组代谢物类别的相对丰度。不同小写字母表示组间差异显著(Tukey HSD检验,p < 0.05)。B:代谢组PLS-DA得分图。ANOVA分析显示:PC1(p = 0.0003);PC2(p = 0.0369)。C:PCA中驱动PC1分离的前20种代谢物的载荷图。D:差异最显著的20种代谢物热图。强度值经log2(x+1)转换后进行颜色映射(蓝色:低;红色:高)。组标签上方的小写字母表示基于Tukey HSD事后检验的组间差异显著性(p < 0.05)。对于每种代谢物,标有不同字母的处理组具有统计学显著差异。E:代谢物添加后的病害发生率。柱状图:均值±标准误(n = 6)。不同字母表示差异显著(ANOVA,Tukey HSD检验;p < 0.05)。LP:低磷处理;NP:正常磷处理;HP:高磷处理。LPDRMs:琥珀酸、壬二酸、苏糖酸、甲硫氨酸;HPDRMs:肌醇、二羟基丙酮、山梨醇、3-氨基异丁酸。LP_HPDRM:低磷基质+HPDRMs;NP_HPDRM:正常磷基质+HPDRMs;HP_LPDRM:高磷基质+LPDRMs;NP_LPDRM:正常磷基质+LPDRMs;LP_CK:低磷+高压灭菌水;NP_CK:正常磷+高压灭菌水;HP_CK:高磷+高压灭菌水。通过整合根际代谢物与微生物群落(真菌和细菌)数据,本研究揭示了磷调控的根际环境与枯萎病易感性差异间的潜在机制。冗余分析(RDA)显示,RDA轴1解释了微生物群落组成31%的变异,其负向与低磷特征根际代谢物(LPDRM)显著关联,表明这些化合物对低磷(LP)组微生物群落具有正向调控作用;RDA轴2独立解释23%的群落变异,其正向主要由高磷特征根际代谢物(HPDRM)驱动,提示这些代谢物对高磷(HP)组微生物群落的积极影响(图4A)。热图展示了标志性微生物类群与特征代谢物的相关性模式。基于欧氏距离和Ward连锁算法的聚类分析发现,特定微生物类群与代谢物形成明显的共现模块:LP组的标志性微生物类群与LPDRM呈强正相关,而与HPDRM呈负相关;反之,HP组相关微生物类群则与其对应的HPDRM化合物呈现强正相关(图4B),这一模式暗示不同磷环境下特定微生物类群与代谢物组可能存在功能关联。采用SparCC相关性算法(|ρ| > 0.7,FDR校正q < 0.05)构建的代谢物-真菌-细菌共现网络,揭示了差异富集真菌、细菌与宿主衍生代谢物间的高阶互作(图4C)。拓扑分析显示所有处理均具有显著模块化结构,其中LP组的功能模块数量显著高于HP组。在LP网络中,有机酸类化合物充当核心枢纽;正常磷(NP)网络的互作以微生物-微生物关联为主;而HP网络则表现出真菌-细菌-代谢物连接性的降低。该网络可视化结果阐明了跨界微生物群落-代谢物互作调控宿主抗病表型的潜在能力。图4. 根际代谢物与微生物群落的整合分析揭示磷依赖性互作。A:微生物群落与代谢物的冗余分析(RDA)。RDA轴1解释31%的微生物群落变异,轴2解释23%的变异。箭头代表代谢物向量。B:特征微生物类群与代谢物的Spearman相关性热图。红色/蓝色分别表示正/负相关(FDR校正p < 0.05)。C:跨界代谢物-微生物共现网络(SparCC算法;|ρ| > 0.7,FDR p < 0.05)。节点大小与中介中心度成正比,蓝色节点为代谢物,红色为细菌,绿色为真菌;红色边表示正相关,蓝色边表示负相关。LP:低磷处理;NP:正常磷处理;HP:高磷处理。为深入探究特定微生物类群及特征代谢物如何参与植物抗病机制,本研究共分离获得288株微生物菌株。其中,低磷(LP)条件下鉴定出代表6个关键属的62株菌株(37株细菌和25株真菌)。通过全面的成对拮抗实验筛选出互容性菌群(即彼此无抑制活性的菌株),最终选取19株细菌和9株真菌进行多维度体外功能表征(图5A)。在两组特征根际代谢物(HPDRM和LPDRM)条件下的生长曲线实验评估了菌株的代谢物利用能力。结果显示:9株测试真菌和19株细菌均无法利用高磷特征代谢物(HPDRM);而部分真菌在LPDRM作为唯一碳氮源的培养基中生长良好,其菌丝生物量显著高于1/5 LB培养基对照组。病原菌(FOC)可高效利用HPDRM,其菌丝产量显著高于对照组,这可能是无菌高磷条件下枯萎病发病率持续高企的原因。绝大多数细菌在LPDRM中的最大生物量显著高于1/5 LB对照组。进一步检测细菌菌株对两组代谢物的趋化响应,发现10株菌对LPDRM表现正向趋化性(图5B,附表8),但部分具备LPDRM利用能力的菌株却未显示趋化响应。特征微生物群对病原菌的拮抗作用被视为植物抗病表型的潜在成因,通过平板对峙实验评估单菌株对FOC的影响,仅5株细菌表现出强拮抗活性(病原菌抑制率>0.4),测试真菌均无直接拮抗作用,但绝大多数真菌菌株表现出对病原菌生长的竞争优势(图5C)。随后评估DRM条件下菌株的土壤定殖效率。7天动态监测显示,LPDRM能显著延长细菌和真菌菌株的土壤定殖时间(图5D、5E)。例如B_3、B_19、F_3、F_4和F_8等菌株在LPDRM环境中培养时,其菌落形成单位(CFU)数量随实验进程持续增加。初始接种量为细菌约10⁶ CFU/g土、真菌10²孢子/g土,在LPDRM辅助下,第7天细菌达10⁷ CFU/g土、真菌达10⁵ CFU/g土的菌株被认定为成功定殖者。图5. 低磷富集微生物分离株的体外功能特性。A:不同培养基中分离株的生长动力学。细菌:最大OD600值;真菌:7天菌丝生物量。对照CK:1/2 LB培养基(细菌)或1/2 PDB培养基(真菌;LPDRM:碳/氮缺陷型无机盐培养基+LPDRM;HPDRM:碳/氮缺陷型无机盐培养基+HPDRM)。B:细菌对代谢物的趋化响应。柱状图:代谢物注射器中的平均细菌计数(n = 6)。LPDRM(100 μM):低磷特征根际代谢物(琥珀酸、壬二酸、苏糖酸、甲硫氨酸);HPDRM(100 μM):高磷特征根际代谢物(肌醇、二羟基丙酮、山梨醇、3-氨基异丁酸);CK:高压灭菌蒸馏水对照;1/5 LB:1/5 LB培养基。C:菌株对病原菌FOC的抑制效应测定。左侧:竞争优势;右侧:抑制率。D-E:添加代谢物的细菌(D)和真菌(E)菌株的土壤定殖动态。曲线表示7天内每克土壤的菌落形成单位(CFU)变化。基于菌株体外互作与定殖能力的综合评估,本研究最终筛选13株细菌与6株真菌构建合成群落(SynCom),以解析其内部互作动态。各菌株的系统发育关系与分类学信息分别见图6A与6B。该SynCom的细菌组分包含6株芽孢杆菌属(Bacillus)、3株贪铜菌属(Cupriavidus)、2株鞘氨醇单胞菌属(Sphingopyxis)及2株德沃斯氏菌属(Devosia);真菌组分为5株曲霉属(Aspergillus)和1株Amesia属。鉴于生物膜在根际微生物定殖与迁移中的关键作用,我们进一步分析了SynCom内细菌-真菌互作关系。结果显示(图6C):(i) 特定菌对存在双向促进效应(表现为细菌生物膜形成与真菌菌落直径同步增加),包括Amesia F7与德沃斯氏菌B2、Amesia F7与芽孢杆菌B10、曲霉F5与芽孢杆菌B18;(ii) 其他组合呈现单向促进模式(一方促进另一方生长但无互惠效应),如曲霉F4与芽孢杆菌B10、曲霉F3与贪铜菌B12、曲霉F5与贪铜菌B12。这些发现揭示了低磷根际微生物生态系统中复杂的真菌-细菌互作网络。为阐明真菌-细菌互作对病原菌拮抗能力的影响,我们开展了SynCom与病原菌的体外拮抗实验。两周后各处理组土壤中病原菌FOC丰度差异显著(图6D)。结果表明:相较于对照,细菌代谢物与真菌活菌联用、真菌代谢物与细菌活菌联用均能显著降低FOC种群数量,且两者效果无显著差异;而完整活菌SynCom的抑菌效果显著优于上述代谢物/活菌组合。最重要的是,LPDRM与SynCom协同处理可使病原菌数量较高磷(HP)对照降低99%,达到最佳抑菌效果。盆栽实验验证了SynCom对黄瓜枯萎病的实际防控效能(图6E,附图8):在HP条件下,单独施用SynCom较HP对照组显著降低发病率50.18%;而HP条件下LPDRM与SynCom协同处理(HP + SynCom + LPDRM)表现出显著协同效应,发病率降低55.1%。这些结果证明,低磷根际条件下形成的微生物-代谢物协同互作机制可在高磷环境中持续建立免疫屏障,有效打破病原菌优势。图6. SynCom-LPDRM协同菌群的构建与生防效能验证。A-B:SynCom菌株的系统发育树(A:13株细菌)与分类组成(B:6株真菌)。C:挥发性有机化合物(VOCs)介导的真菌-细菌互作。数据经相应单菌株对照标准化处理。D:SynCom与代谢物处理后土壤中病原菌(FOC)的丰度变化。柱状图:均值±标准误(n=6)。不同字母表示差异显著(ANOVA,Tukey HSD检验;p<0.05)。E:高磷(HP)土壤中添加SynCom±LPDRM对黄瓜病害发生率的影响。柱状图:均值±标准误(n=6)。不同字母表示差异显著(ANOVA,Tukey HSD检验;p<0.05)。LPDRM:低磷特征根际代谢物(琥珀酸、壬二酸、苏糖酸、甲硫氨酸);HPDRM:高磷特征根际代谢物(肌醇、二羟基丙酮、山梨醇、3-氨基异丁酸);BM+fungi:细菌代谢物+真菌悬浮液;FM+bacteria:真菌代谢物+细菌悬浮液;SynCom:完整微生物悬浮液;SynCom+LPDRM:完整微生物悬浮液+低磷特征代谢物;HP+Syn:高磷处理+完整微生物悬浮液;HP+Syn+LPDRM:高磷处理+完整微生物悬浮液+低磷特征代谢物。本研究揭示了磷素依赖性调控根际微生物群落组装与土传病害易感性的机制。实验证实:低磷有效性促进形成以有机酸为核心的随机组装型稳定微生物-代谢物网络,从而增强对枯萎病的抗性;而高磷条件则驱动确定性组装并富集促进病原菌增殖的代谢物,进而提高植物病害易感性(图7)。该研究将磷素管理的视角从单一关注植物营养拓展至统筹土壤生态健康与病害动态,表明长期磷累积的农业系统面临更高的作物病害风险。因此,合理施磷策略需兼顾磷素对土壤微生物群落生态和植物营养的双重效应。在可持续农业框架下,未来研究应进一步探索不同作物类型与病害的磷驱动机制,以及如何通过优化磷管理来协同提升土壤健康与作物抗病性。