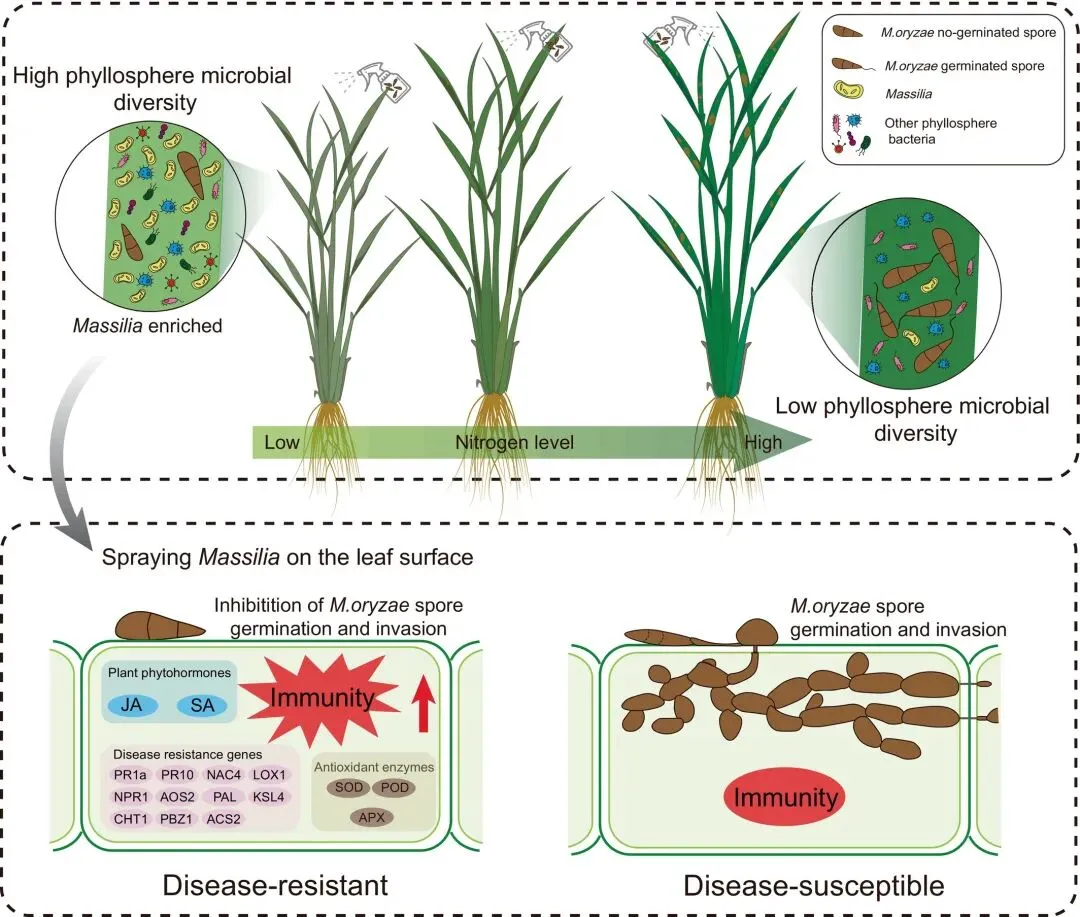
1. 高氮施用加剧田间与温室环境下的稻瘟病症状
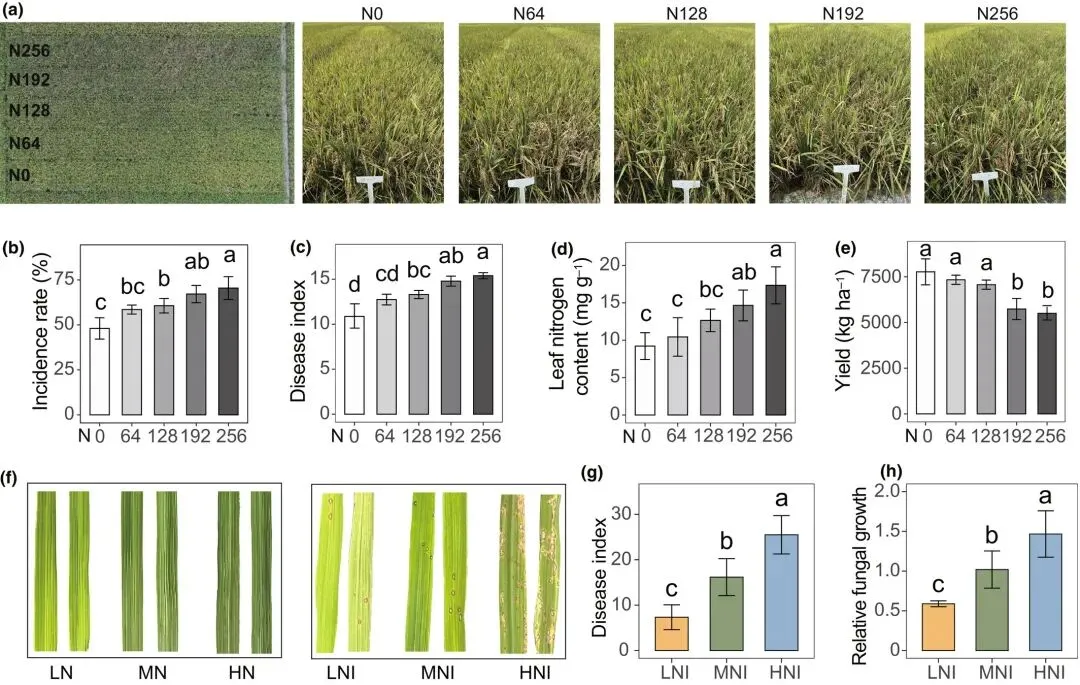
为探究不同氮水平对水稻健康的影响,我们开展了一项田间调查,旨在量化不同施氮量下稻瘟病的发生情况。低氮条件下,水稻植株稀疏、矮小且叶片偏黄;而高氮条件则促进植株高大、叶片深绿、群体茂盛。尽管外观旺盛,但田间观察发现高氮供应的植株稻瘟病病斑更严重且更易倒伏,而低氮供应植株病斑较少且保持直立(图1a)。随着施氮量增加,稻瘟病发病率、病害指数及叶片氮含量均逐渐上升(图1b-d)。具体而言,在256 kg·ha⁻¹的施氮水平下,稻瘟病发病率达峰值70.45%(图1b),同时水稻产量显著低于低氮处理(图1e)。
图1 施氮对稻瘟病发生与严重程度的影响。(a)不同施氮量(N0、N64、N128、N192、N256分别表示0、64、128、192、256 kg·N·ha⁻¹)下感染稻瘟病菌(Magnaporthe oryzae)稻田的俯视图与侧视图。(b-e)田间不同氮处理下的稻瘟病发病率(b)、稻瘟病病害指数(c)、叶片氮含量(d)及水稻产量(e)。(f)温室实验中,1 mM N(低氮)、2 mM N(中氮)或5 mM N(高氮)处理下未接种或接种稻瘟病菌的水稻叶片表型。(g-h)温室实验中的稻瘟病病害指数(g)和病叶中病原菌相对生长量(h)。数据为均值±标准差(n=6)。不同字母表示Tukey检验下的显著差异(P < 0.05)。
为更详细探究氮水平与稻瘟病的关系,我们在可控条件更强的温室中开展实验。对三种不同供氮水平下生长的水稻接种稻瘟病菌(图1f)。与田间调查一致,氮水平升高显著促进稻瘟病发展并提高病原菌丰度(图1g,h)。综上结果表明,氮肥施用与稻瘟病严重程度呈正相关。
2. 氮水平与稻瘟病菌侵染影响水稻叶际微生物组
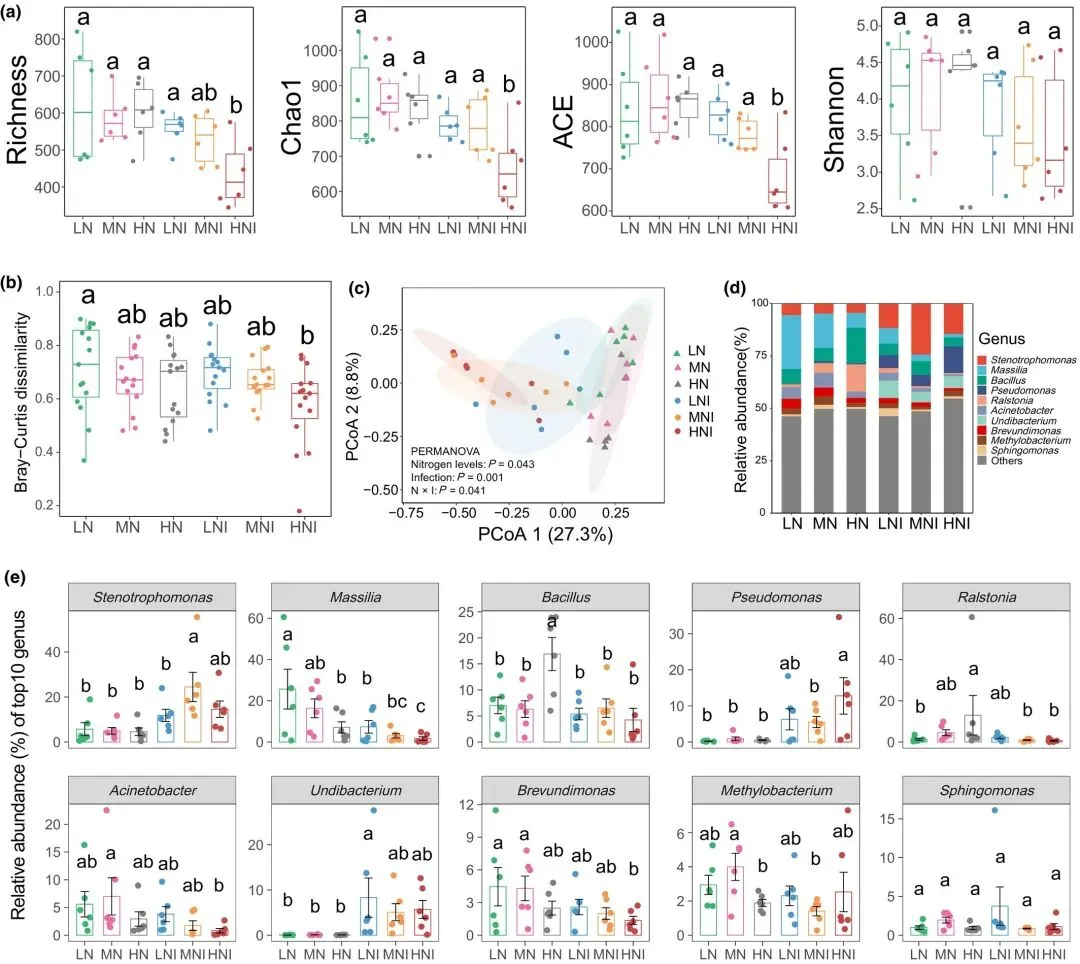
鉴于叶际微生物在病原抗性中的作用,我们探究了氮供应如何影响健康与感病水稻植株叶片微生物群落的组成。经过质量过滤,我们从36份叶片样本中保留了2,741,161条高质量细菌序列用于下游群落组成分析。Alpha多样性指数(丰富度、ACE和Chao1)显示,接种病原菌的低氮(LNI)与高氮(HNI)处理组植株叶际细菌群落存在显著差异(图2a)。基于Bray-Curtis相异矩阵和PERMANOVA的PCoA分析表明,氮水平、病原菌接种及其交互作用均显著影响叶际细菌群落组成(图2b,c)。无论稻瘟病菌侵染状态如何,低氮条件下Massilia菌属的相对丰度均显著高于高氮条件(图2d,e)。这些结果说明氮素有效性与病原侵染共同塑造水稻叶际微生物群的多样性和组成,进而潜在影响植株健康状态。
图2 氮水平与稻瘟病菌接种对水稻叶片内生细菌多样性及组成的影响。(a)不同氮处理与侵染条件下水稻叶片内生细菌的Alpha多样性指标:丰富度、Chao1、基于丰度的覆盖度估计值(ACE)和香农指数。(b)不同氮水平与侵染条件下内生细菌群落的Bray-Curtis距离。(c)基于Bray-Curtis距离的所有叶片样本内生细菌群落主坐标分析图。(d)属水平细菌群落的相对丰度。(e)所有叶片样本中前10位内生细菌属的相对丰度。数据为均值±标准差(n=6)。不同字母表示Tukey检验下的显著差异(P < 0.05)。通过置换多元方差分析(PERMANOVA)判定不同氮水平、侵染条件及其交互作用对细菌β多样性的影响。LN:低氮;MN:中氮;HN:高氮(未接种稻瘟病菌);LNI、MNI、HNI分别表示不同氮水平下接种稻瘟病菌的处理。箱线图显示第一和第三四分位数,箱内横线为中位数,须线为不超过箱体四分位距1.5倍的极值点。
3. 潜在关键叶际细菌赋予水稻对稻瘟病的抗性
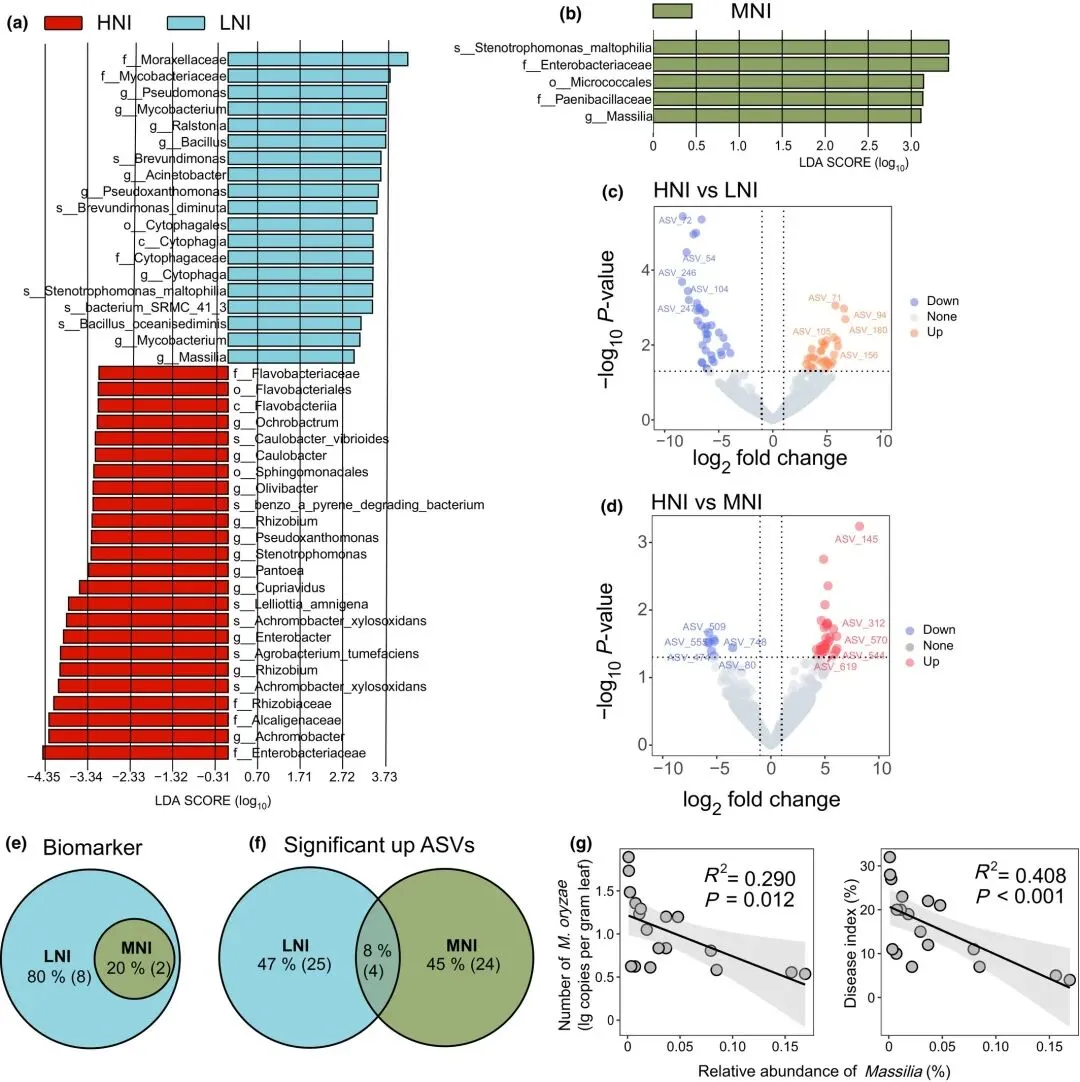
LEfSe方法分别鉴定了低氮接种(LNI)与高氮接种(HNI)样本间(图3a;表S7),以及中氮接种(MNI)与高氮接种(HNI)样本间(图3b;表S7)的差异性细菌生物标志物。Massilia(马赛菌属)和Stenotrophomonas(窄食单胞菌属)是LNI与MNI样本叶际微生物组中仅共享的两个生物标志物(图3e)。采用DESeq2进行的差异丰度分析识别出HNI与LNI样本间(图3c;表S8)以及HNI与MNI样本间(图3d;表S8)显著上调的扩增子序列变体(ASVs)。四个ASV(ASV 178、ASV 11、ASV 148和ASV 156)在LNI和MNI样本中的丰度均持续高于HNI样本(图3f),其中ASV 148被分类属于Massilia属。相反,五个ASV(ASV 509、ASV 606、ASV 555、ASV 474和ASV 808)在LNI和MNI样本中的丰度均持续低于HNI样本(图S2),这些ASV包括来自Novosphingobium属(ASV 509)、Acidovorax属(ASV 606)和Pseudomonas属(ASV 474)的分类单元。
图3 不同氮处理下稻瘟病菌侵染相关潜在细菌类群的鉴定。(a, b)线性判别分析效应量(LEfSe)(对数LDA值>3,Wilcoxon P<0.05)显示分别将抗病组LNI(a)和MNI(b)与感病组HNI进行比较时最显著的生物标志物。下划线前的小写字母表示分类等级:c,纲;f,科;g,属;o,目;s,种。(c, d)火山图展示抑制组LNI(c)和MNI(d)与非抑制组HNI相比富集和耗竭的扩增子序列变体(ASVs)。(e, f)韦恩图显示LNI和MNI样本间共享的生物标志物和富集的ASVs。(g)探索Massilia相对丰度与水稻叶片中稻瘟病菌丰度(左)及病害指数(右)的相关性。散点图显示合并数据、最佳拟合线(实线)及其95%置信区间(灰色阴影)。LNI、MNI和HNI分别表示接种稻瘟病菌的低氮、中氮和高氮处理。
基于前述分析,我们确定Massilia是参与抗病性的关键叶际微生物。在LEfSe和DESeq2分析中,与高氮感病植株(HNI)相比,其在低氮或中氮抗病植株(LNI和MNI)中持续富集。此外,我们检测到Massilia的相对丰度与水稻叶片中稻瘟病菌丰度(R² = 0.290, P = 0.012)及稻瘟病病害指数(R² = 0.408, P < 0.01)均呈负相关(图3g)。
4. 马赛菌(Massilia)抑制稻瘟病
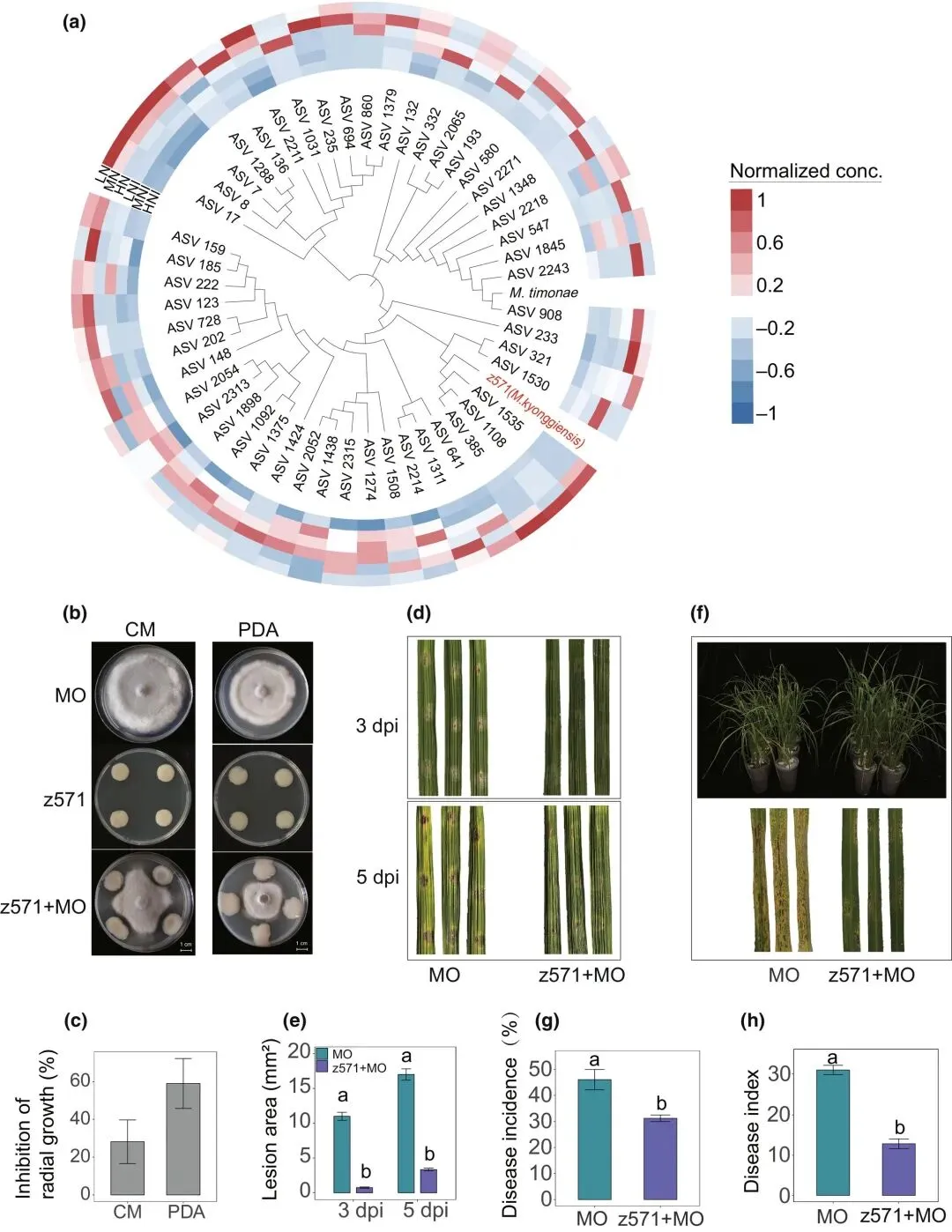
通过序列比对(表S3)和基于16S rDNA序列的系统发育分析(图4a),我们确定从江苏省农业微生物种质资源库获得的细菌分离株z571属于马赛菌属(Massilia)。对峙培养试验表明,马赛菌z571菌株在完全培养基(CM)和马铃薯葡萄糖琼脂(PDA)平板上均可抑制稻瘟病菌(M. oryzae)的生长(图4b,c)。
图4 马赛菌对稻瘟病菌拮抗作用的验证。(a)基于邻接法构建的系统发育树,显示叶片样本中检测到的马赛菌扩增子序列变体(ASVs)、参考菌株Massilia timonae及马赛菌分离株z571的进化关系。(b)z571在完全培养基和PDA平板上对稻瘟病菌径向生长的抑制效果。MO:仅接种稻瘟病菌;z571:仅接种马赛菌;z571+MO:共接种z571菌株与稻瘟病菌。(c)各培养基上真菌径向生长的抑制率(对应图b)。(d)离体水稻叶片接种稻瘟病菌(MO)或联合接种z571(z571+MO)后3天与5天(dpi)的病斑代表图。(e)离体水稻叶片接种稻瘟病菌后3天与5天的病斑面积(对应图d)。(f)水稻幼苗生长表型(上图)及叶片病害症状(下图):MO处理与z571+MO联合处理对比。(g,h)z571对水稻叶片发病率(g)和病害指数(h)的影响。数据为均值±标准差(n=6)。不同字母表示Tukey最小显著差异法检验的显著差异(P < 0.05)。
因此,我们进一步探究了分离株z571在体外和体内对稻瘟病的抑制能力。在离体水稻叶片接种稻瘟病菌前施用z571菌悬液,可显著减小病斑面积并限制病斑扩展(3 dpi与5 dpi)(图4d,e)。相比之下,仅喷洒无菌水对照的叶片形成 elongated坏死病斑(图4d)。温室实验中,向水稻植株喷施z571菌悬液显著降低了接种稻瘟病菌后的发病率和病害指数(图4f-h)。此外,z571处理还促进植株生长:与清水对照相比,喷施z571的植株株型更挺立、叶色更深绿(图4f),表现为生物量、株高和SPAD值均显著增加(图S3)。这些结果表明,马赛菌z571可作为稻瘟病的生防菌剂,同时增强植物抗性并促进水稻生长。
5. 马赛菌控制稻瘟病的第一种机制:直接拮抗病原菌发育
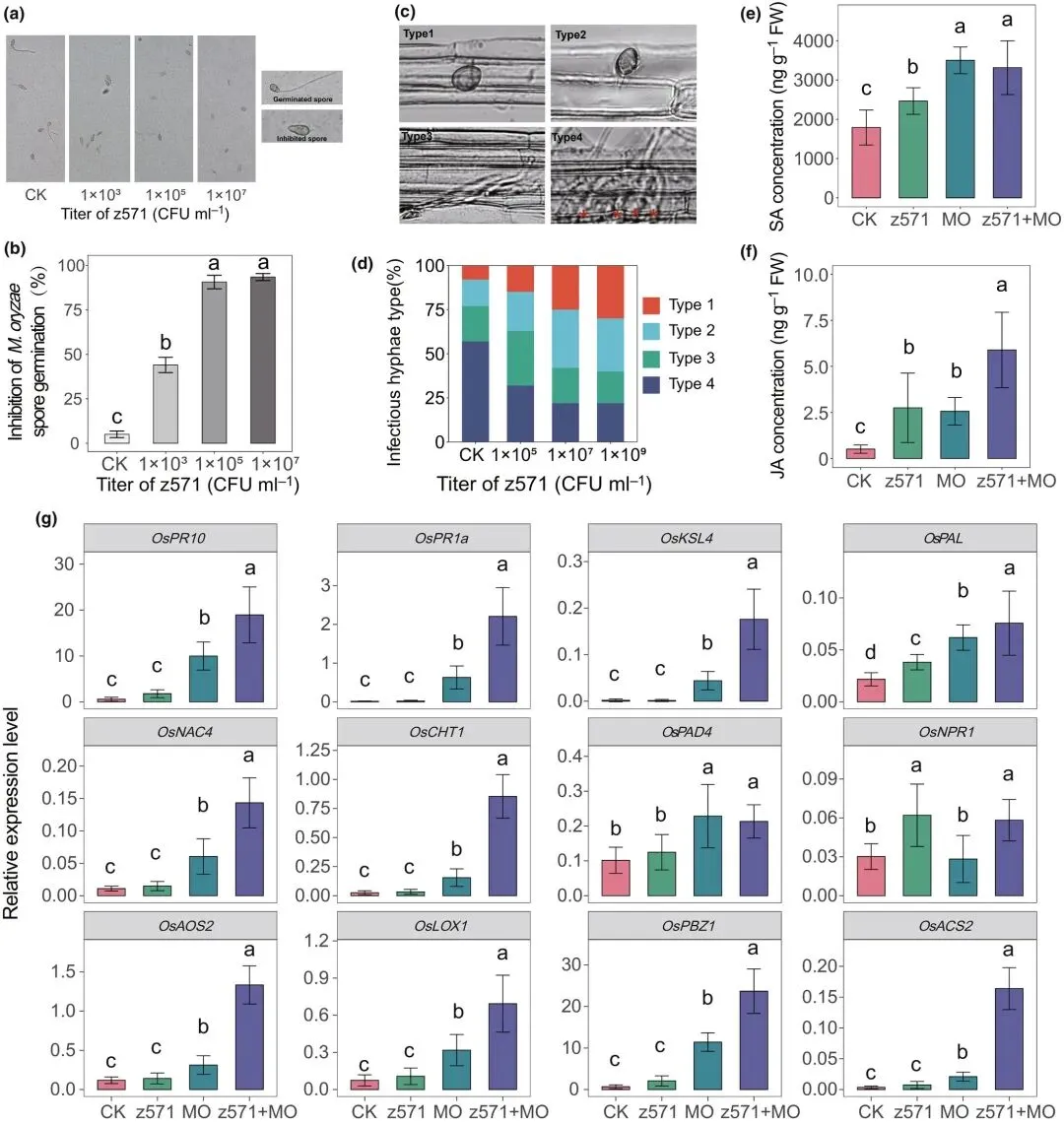
研究发现,当与稻瘟病菌分生孢子悬液共培养时,马赛菌z571能显著抑制孢子萌发。显微观察表明,随着细菌添加量的增加,z571对稻瘟病菌孢子萌发的抑制效果增强。当菌悬液浓度为1×10⁵ CFU·ml⁻¹时,孢子萌发几乎被完全抑制(图5a,b)。
图5 马赛菌介导的稻瘟病菌抑制机制。(a)光镜下观察z571悬液对体外稻瘟病菌孢子萌发的影响。(b)不同浓度z571菌悬液对稻瘟病菌孢子萌发的抑制率。(c)水稻鞘细胞内侵染性稻瘟病菌菌丝的评估,分为四种类型。(d)不同z571浓度下100个附着胞侵染点中各类侵染性菌丝的百分比。(a、b、d)中CK=无菌水处理稻瘟病菌(未加z571)。(e-g)不同接种处理下水稻叶片的水杨酸含量(e)、茉莉酸含量(f)及防御基因相对表达量(g)(CK:仅无菌水处理;z571:仅接种z571菌株;MO:仅接种稻瘟病菌;z571+MO:共接种z571菌株与稻瘟病菌)。数据为均值±标准差(n=6)。不同字母表示Tukey最小显著差异法检验的显著差异(P < 0.05)。
对接种稻瘟病菌48小时后水稻鞘细胞内侵染性菌丝生长的分析表明,随着z571菌悬液浓度的升高,Ⅲ型(扩展但限于单个细胞)和Ⅳ型(扩展至多个细胞)侵染性菌丝的比例下降。当菌悬液浓度为1×10⁷ CFU·ml⁻¹时,z571处理对菌丝侵染的抑制效果达到最大,仅约38%的附着胞侵染点被归类为Ⅲ型或Ⅳ型侵染性菌丝。相比之下,清水处理的水稻鞘细胞表现出更高水平的侵染,75%以上的侵染点存在Ⅲ型和Ⅳ型菌丝(图5c,d)。
6. 马赛菌控制稻瘟病的第二种机制:通过防御预激活诱导宿主抗性
除直接抑制作用外,z571还能增强宿主对稻瘟病菌的免疫响应。与未处理叶片相比,z571处理导致水稻叶片中水杨酸(SA)和茉莉酸(JA)浓度升高。特别值得注意的是,接种稻瘟病菌后,经z571处理的植株JA水平显著高于单独接种稻瘟病菌的植株(图5e,f)。此外,在稻瘟病菌侵染后,与单独接种稻瘟病菌的植株相比,z571处理植株中多个防御相关基因的表达水平显著上调,包括病原相关基因(OsPR10、OsPR1a、OsKSL4、OsPAL、OsNAC4和OsACS2)、SA信号标记基因(OsCHT1和OsNPR1)以及JA标记基因(OsAOS2、OsLOX1和OsPBZ1)(图5g)。
与前述结果一致,喷施z571并接种稻瘟病菌的植株中抗氧化酶(SOD、POD和APX)活性显著高于单独接种稻瘟病菌的植株(图S4)。综上结果表明,马赛菌z571通过抑制病原菌生长并预激活多层次的宿主免疫系统,从而增强对稻瘟病的抗性。
7. 马赛菌(Massilia)是对多种病原菌的潜在生防因子
我们评估了马赛菌在琼脂平板上对几种重要作物病原菌的拮抗潜力。马赛菌强烈抑制以下病原菌的生长:Fusarium fujikuroi(藤仓镰刀菌,图S5b)、Fusarium graminearum(禾谷镰刀菌,图S5c,e)、Nigrospora oryzae(稻黑孢菌,图S5d)、Ustilaginoidea virens(稻曲病菌,图S5f)、Phytophthora capsici(辣椒疫霉,图S5g)和Sarocladium oryzae(稻帚枝霉,图S5h)。这些病原菌可侵染多种禾本科作物并引发辣椒(Capsicum spp.)病害。在所有测试中,马赛菌均形成清晰抑菌圈,抑制生长率介于15%至90%之间(图S5i),凸显其广谱生防活性。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
