1区top(IF:12.5)│南京农大:薤白多糖通过调控肠道微生物和代谢缓解溃疡性结肠炎
- 2026-05-18 09:49:43
 点击蓝字“代谢组metabolome”,轻松关注不迷路
点击蓝字“代谢组metabolome”,轻松关注不迷路

生科云网址:https://www.bioincloud.tech/
编译:微科盟 草重木雪,编辑:微科盟X、江舜尧。
微科盟原创微文,欢迎转发转载。
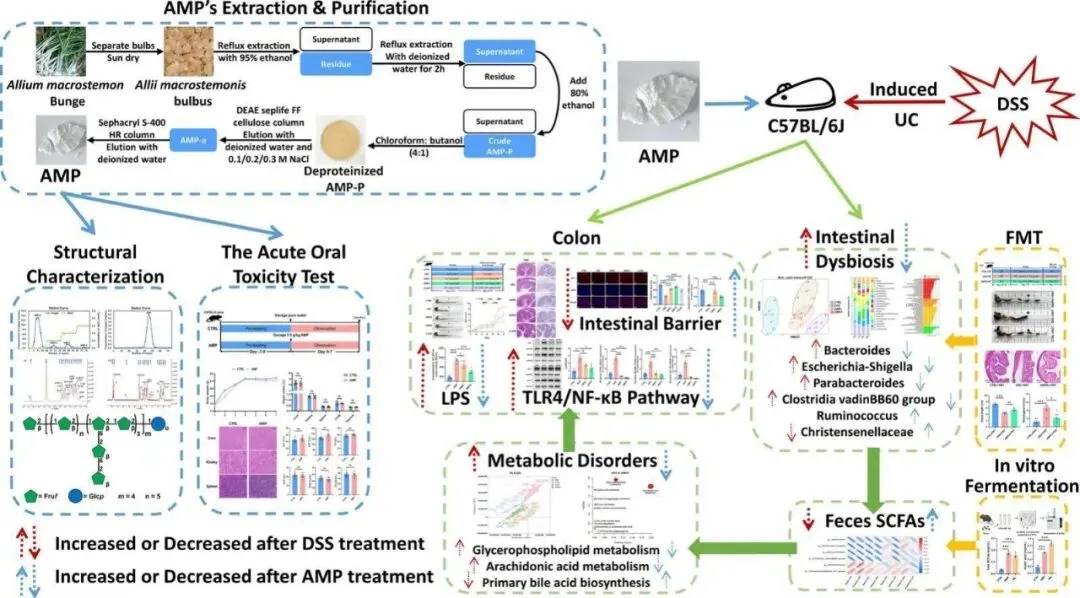
论文ID
IF:12.5
DOI号:10.1016/j.carbpol.2026.125174
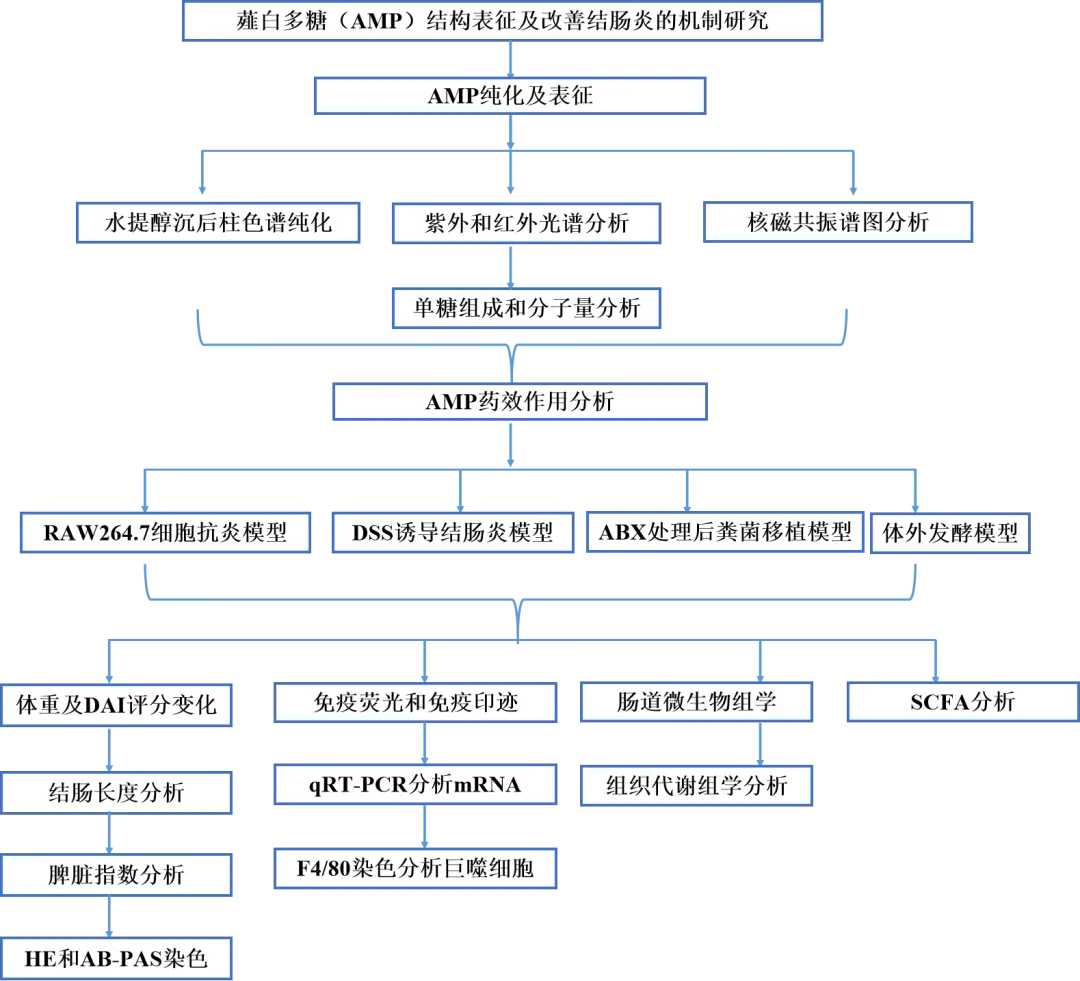
实验设计
结果
1. AMP的分离、纯化和基本特征
研究者通过热水提取和乙醇沉淀获得粗多糖(AMP单键P)(图1A)。然后将AMP-P加入DEAE-Sepharose FF柱中,用不同浓度的NaCl溶液洗脱四个组分。其中,从管7至19洗脱的AMP-a是最丰富的产物,占总收率的83.50%(图1B)。主要组分(AMP-a)通过Sephacryl S-400HR柱进行额外纯化,得到精制的多糖AMP(图1C)。在紫外光谱中,AMP在280和260 nm处没有吸收,表明它具有高纯度(图1D)。AMP的红外光谱(图1E)在3386 cm-1处显示了一个广泛分散的强峰,表明O-H和2934.81 cm-1处的C-H带,表明AMP具有多糖的吸收峰特征。在1638.02 cm-1附近的相对较强的吸收峰表明了C=O双键的特征。多糖在950–1200 cm-1范围内的吸光度是发现C-O-C和C-O-H连接带位置的地方。931.60 cm-1和814.27 cm-1的吸收带与带有β-糖苷键的呋喃糖环结构一致。350-600cm-1范围内的谱带被指定为吡喃糖环的骨架模式。AMP的单糖组成为95.99%的Fru和3.69%的Glc(图1F)。表1列出了AMP的摩尔质量分布和单糖组成的摩尔质量百分比。SEC-MALLS-RI结果表明,AMP的分子量为9.007 kDa。AMP的均匀性由1.33的分散度值证实,在绝对分子量色谱图中检测到单个对称峰(图1G)。
表1AMP单糖组成的分子量和摩尔质量百分比。

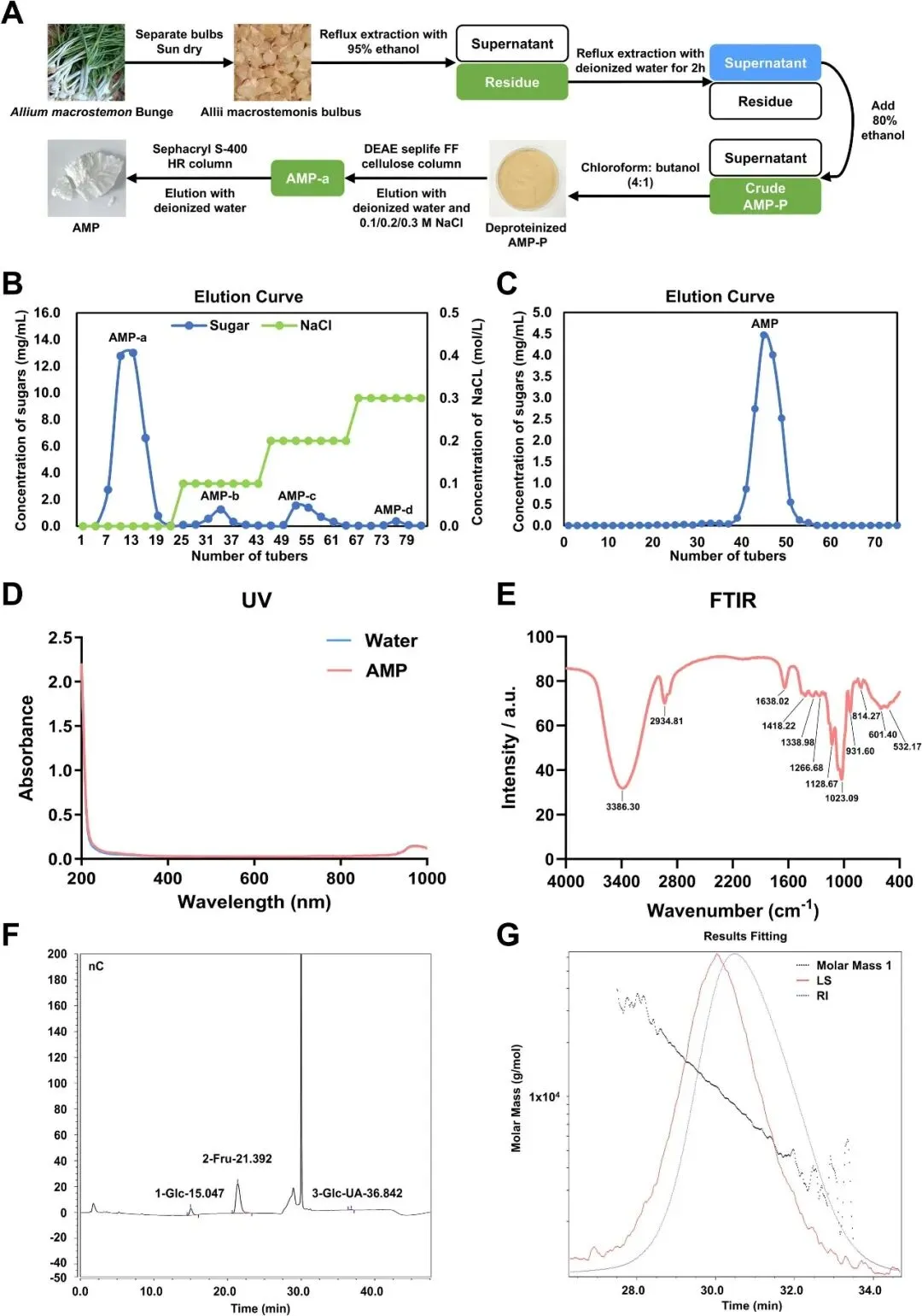
图1 AMP的分离、纯化过程和基本特征。(A)提取和纯化程序。(B)通过离子交换色谱法对AMP-P的洗脱曲线。(C)AMP-a的凝胶过滤色谱洗脱模式。(D)AMP多糖组分的紫外光谱。(E)AMP多糖组分的FTIR光谱。(F)AMP单糖组成离子色谱图。(G)AMP的绝对分子量色谱图。
2. AMP的结构表征
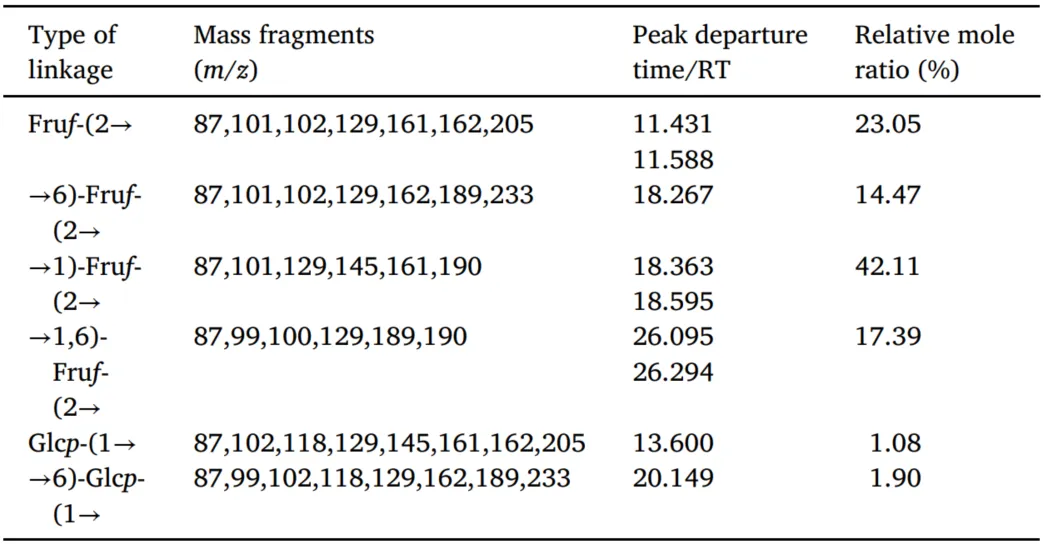
甲基化分析为单糖的连接位置和分支模式提供了关键见解,从而促进了AMP的结构阐明。与参考数据库的比较显示,AMP主要由六个糖苷残基组成(图S2),详细结果总结在表2中。Fru残基表现出四种不同的连接类型:Fruf-(2→,→6)-Fruf-(2→、→1)-Fruf-(2→和→1,6)-Fruf-(2→。基于峰面积的定量分析表明→1)-Fruf-(2→是主要的连接(42.11%),其次是 Fruf-(2 →(23.05%), →1,6)-Fruf-(2 → (17.39%),以及→6)-Fruf-(2 →(14.47%)。→1,6)-Fruf-(2 →清楚地表明AMP是一种支链多糖。鉴于Glc所占比例最低,它可能不参与AMP的主要重复单元,而是作为连接到多糖骨架的末端残基存在。总之,本研究结果表明AMP是一种支化聚合物,其主要特征是中心重复结构中的Fru,而Glc位于主链的末端位置。
表2AMP键合结构分析结果。
核磁共振波谱研究用于进一步阐明AMP的糖苷连接。NMR光谱和相应的化学位移分配如图2A-F所示。化学位移值与甲基化分析结果一致,组合结构信息如表3所示。
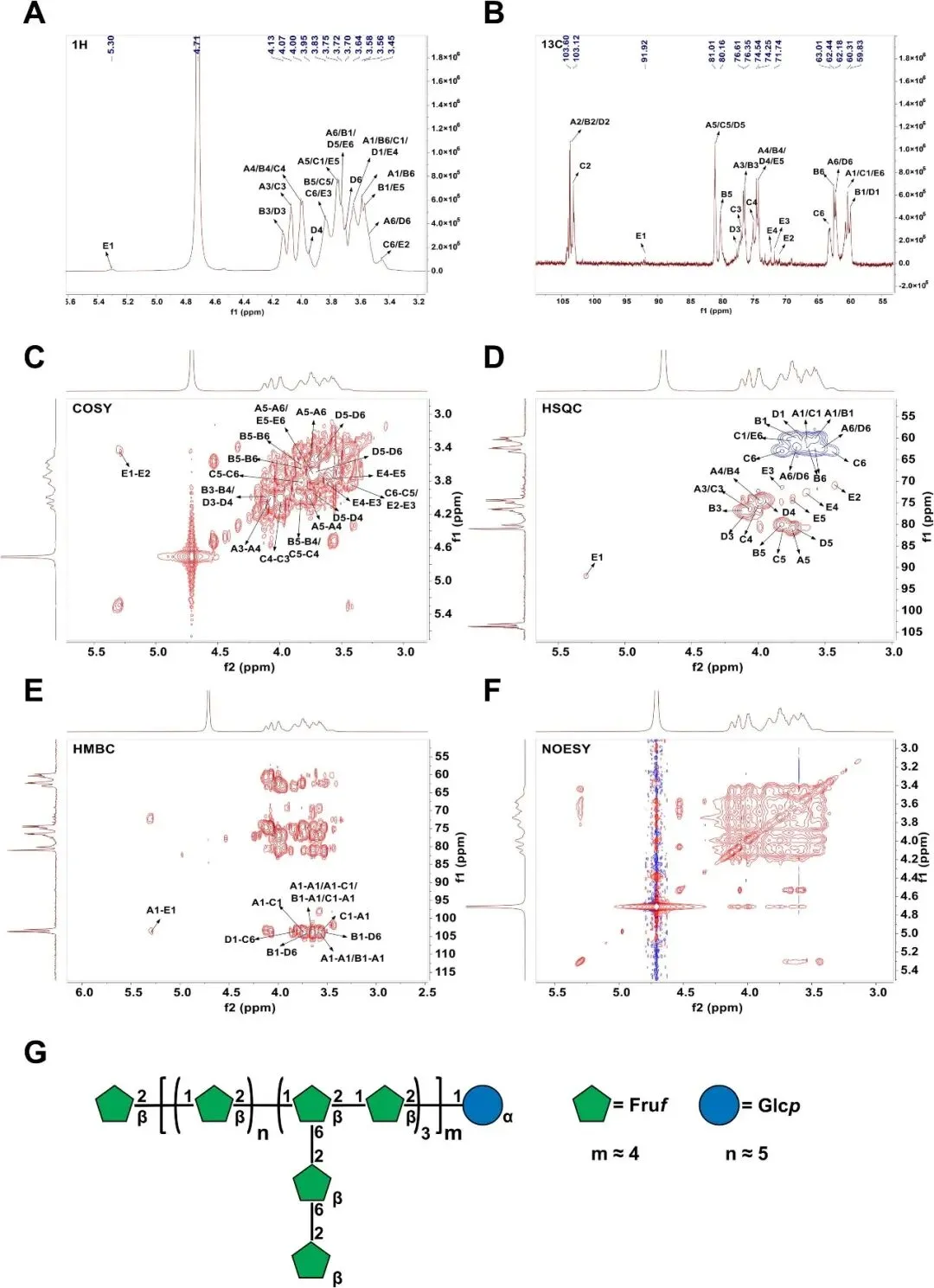
图2 AMP的结构特征。(A)AMP的1H、(B)13C、(C)COSY、(D)HSQC、(E)HMBC和(F)NOESY谱。(G)AMP推断结构示意图。
表3 AMP的1H和13C NMR化学位移。
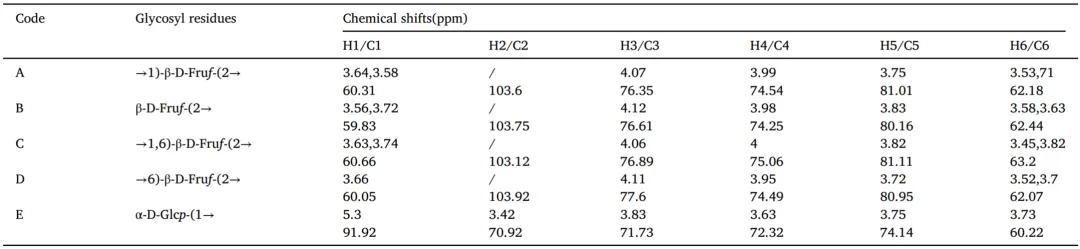
AMP的13C NMR分析揭示了δ103.6、103.75、103.12和103.92 ppm处的四个不同异头碳信号,这些信号归因于糖残基A至D。1H NMR谱在4.4-5.3 ppm范围内没有显示可观察到的峰,HSQC谱在异头区域缺乏这些残基的交叉峰,这清楚地证明了Fru构象。结合HSQC相关性和甲基化结果,残基的特征如下:(A)→1)-β-D-Fruf-(2→,(B)β-D-Fruf-(2→,(C) →1,6)-β-D-Fruf-(2→,和(D)→6)-β-D-Fruf-(2→。在两个谱中,δ5.3/91.92ppm(1H/13C)处出现了一个单独的异头信号,称为E。鉴于糖苷键中的α-异头碳通常出现在δ95-100ppm附近,E被确认为α-d-葡萄糖单元。COSY数据显示,E的H2质子在δ3.42处,而H3-H6分别在δ3.83、3.63、3.75和3.73ppm处共振。HSQC进一步验证了E在δ91.92、70.92、71.73、72.32、74.14和60.22 ppm处从C1到C6的碳转移,与O-1取代一致。通过与先前研究的交叉引用甲基化分析,E被确定为α-D-Glcp-(1→)。
由于在NOESY谱中没有检测到相关性,因此AMP的连接模式主要使用HMBC谱确定。关键的核磁共振互相关注释如下。对于残基A,C2与A的H1(δ103.6/3.58-3.64ppm)和C的H1(Δ103.6/3.63-3.74ppm)接触。残基B表现出与A的C2-H1连接(δ103.75/3.58-3.64 ppm)和与D的C2-H6相关性(δ103.95/3.52-3.70 ppm)。进一步的连接包括C的C2到A的H1(δ103.12/3.58-3.64 ppm)和D的C2到C的H6(δ103.92/3.82 ppm)。
基于全面的结构分析,AMP主要由以下核心结构组成:→1)-β-D-Fruf-(2→和→1,6)-β-D-Fruf-(2→连接。此外,β-D-Fruf-(2→6)-β-D-Fruf-(2→残基连接到→1,6)-β-D-Fruf-(2→的O-6位置,形成短侧链。图2G提供了推断AMP架构的详细示意图。
3. AMP改善DSS诱导的小鼠UC症状
RAW264.7细胞炎症模型是一种常见的体外抗炎功效筛选模型。因此,研究者在药物筛选的早期阶段使用了它。CCK-8测定显示,AMP在150μg/mL以下不影响RAW264.7细胞的正常生长(图3A)。因此,研究者用AMP(50或100μg/mL)处理LPS刺激的RAW264.7细胞,然后提取总RNA以评估炎症相关因子的转录变化。RT-qPCR分析显示,AMP剂量依赖性地降低了LPS诱导的IL-1β、IL-6和TNF-α的上调(图3B-D),表明其在LPS刺激的RAW264.7巨噬细胞中具有强大的抗炎作用。此外,急性经口毒性研究表明,AMP作为功能性食品具有很高的安全性和潜力(图S2)。
基于AMP抗炎活性的体外研究结果和AMP的高安全性,研究者研究了其在小鼠中的治疗效果。如图3E所示,研究者使用DSS在小鼠中诱导结肠炎。在整个研究过程中,AMP治疗缓解了DSS相关的体重和DAI变化(图3F-G)。实验结束时,组织采集显示AMP显著改善了DSS诱导的结肠缩短和脾脏增大(图3H-K)。结肠组织的组织病理学分析表明,AMP减轻了DSS诱导的炎性细胞浸润、隐窝损伤和上皮脱离,同时也逆转了DSS诱导杯状细胞数量的减少(图3L-N)。总体而言,数据表明AMP显著减轻了DSS诱导的小鼠结肠炎,低剂量显示出与5-ASA相似的效果,高剂量显示出更优的效果。
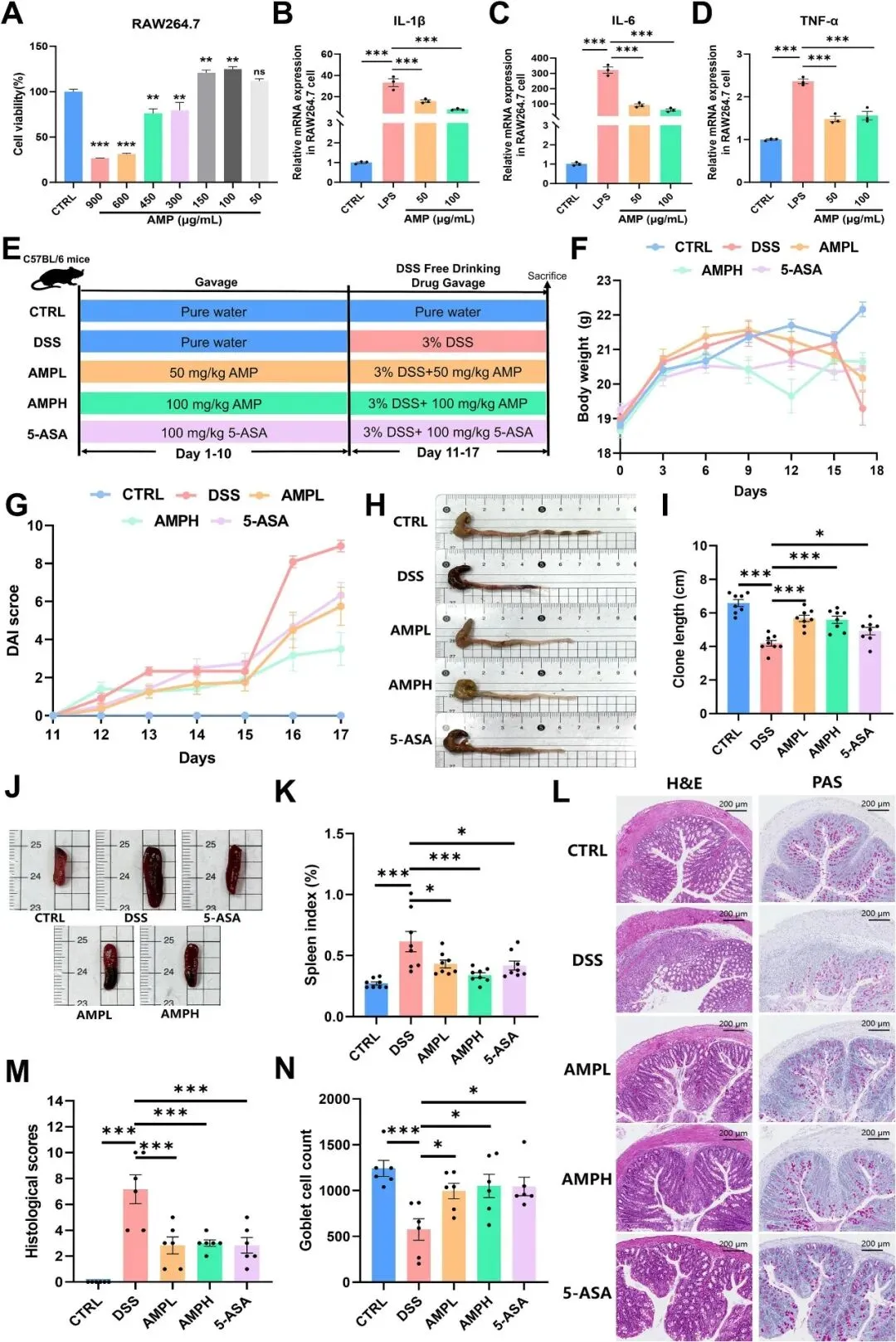
图3 AMP在体外可以抑制DSS诱导的小鼠的细胞炎症,缓解UC症状。(A)暴露于LPS的24小时AMP处理后的RAW264.7细胞存活率;n=3。RAW264.7细胞中IL-1β(B)、IL-6(C)和TNF-α(D)的(B-D)mRNA水平;n=3。(E)动物实验示意图。(F)体重变化曲线,(G)DAI评分,(H)结肠取样形态,(I)结肠长度,(J)脾脏取样形态,以及(K)脾脏指数;n=8。(L)左:H&E染色;右图:每组小鼠结肠的PAS染色图像。(M)组织学评分和(N)杯状细胞计数;n=6。
4. AMP恢复UC小鼠的肠屏障
DSS破坏肠道屏障并诱导小鼠结肠炎,研究者因此评估AMP对屏障完整性的影响。免疫荧光分析表明,DSS导致关键屏障蛋白(ZO-1、Occludin和Claudin)水平降低,而AMP有效地恢复了它们的表达(图4A-F)。一致的是,RT-qPCR分析显示AMP显著增加了ZO-1、Occludin、Claudin和MUC2的mRNA表达(图4G-J)。总体而言,结果表明,AMP显著增强了DSS小鼠中紧密连接蛋白的表达和转录,AMPL和AMPH干预效果接近5-ASA。
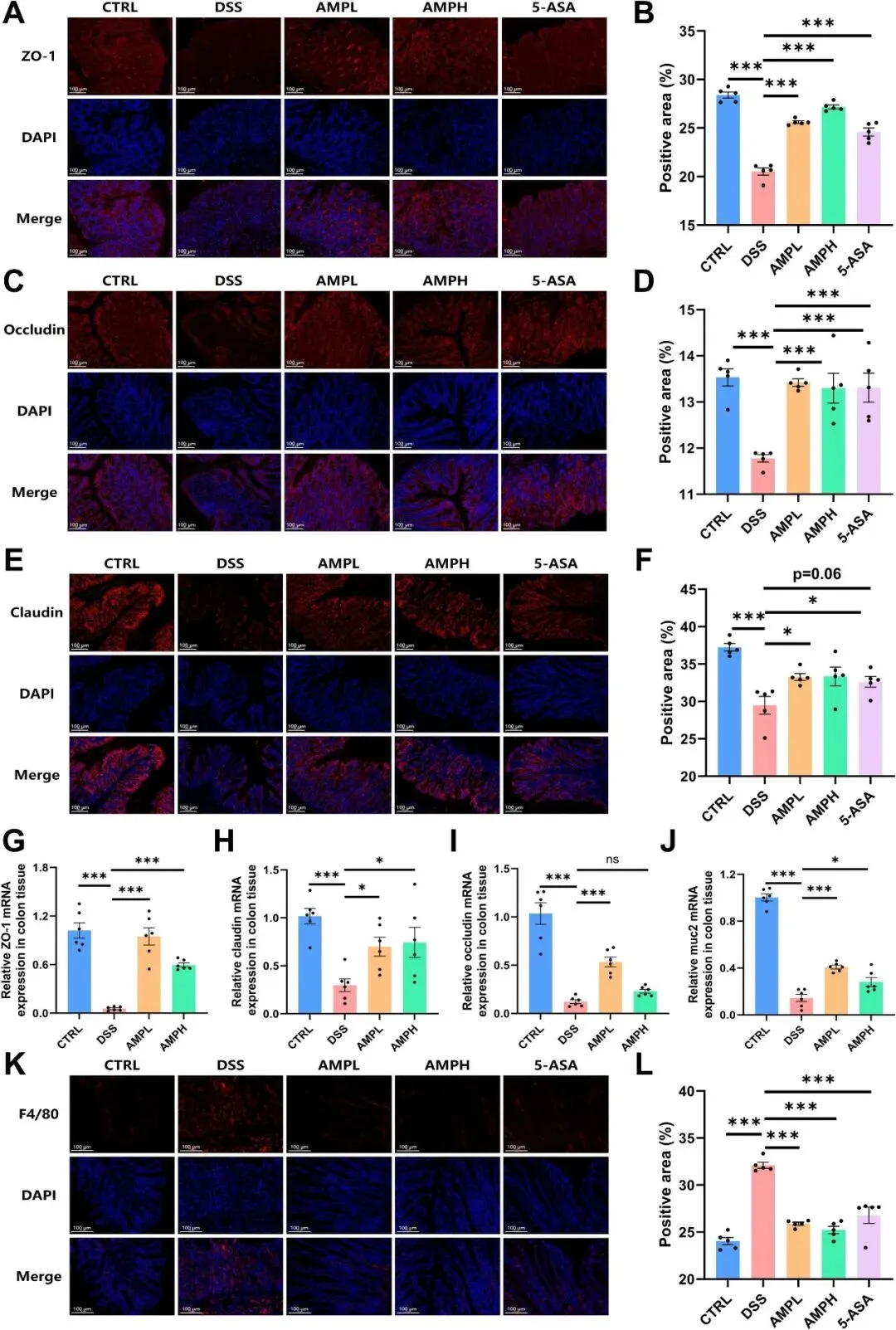
图4 AMP可恢复UC小鼠的肠道屏障。(A-F)来自ZO-1(A)、Occludin(C)和Claudin(E)免疫荧光的选择性图像;ZO-1(B)、Occludin(D)和Claudin(F)免疫荧光染色的强度分析;n=5。结肠组织中ZO-1(G)、Claudin(H)、Occludin(I)和MUC2(J)的mRNA水平;n=6。(K-L)F4/80免疫荧光染色的选择性图像(K)和强度分析(L);n=5。
5. AMP通过减少LPS侵袭和抑制TLR4/NF-κB通路减少DSS小鼠的肠道炎症
在小鼠模型中,DSS诱导的UC是由肠道屏障减弱引起的,允许LPS进入结肠组织并激活TLR4/NF-κB信号级联,从而引发组织炎症。F4/80免疫荧光染色结果显示,DSS处理诱导结肠中巨噬细胞大量积聚,而AMP处理显著减弱了DSS驱动的浸润(图4K-L)。DSS给药提高了循环IL-1β、IL-6和TNF-α,而AMP干预则剂量依赖性地减轻了这些DSS诱导的不良反应(图5A-C)。与所有其他组相比,DSS处理的小鼠结肠LPS丰度升高(图5D)。接下来,研究者定量了结肠组织中IL-1β、IL-6和TNF-α的转录和蛋白质丰度。AMP显著降低了结肠炎症细胞因子的输出(图5E-J)。此外,与CTRL相比,DSS显著激活了TLR4/NF-κB通路关键蛋白水平,并提高了p65磷酸化(pp65/p65比值),而AMP干预有效地逆转了这些变化(图5K-R)。总的来说,这些结果表明AMP通过减少LPS易位和TLR4/NF-κB信号通路来减轻组织炎症。
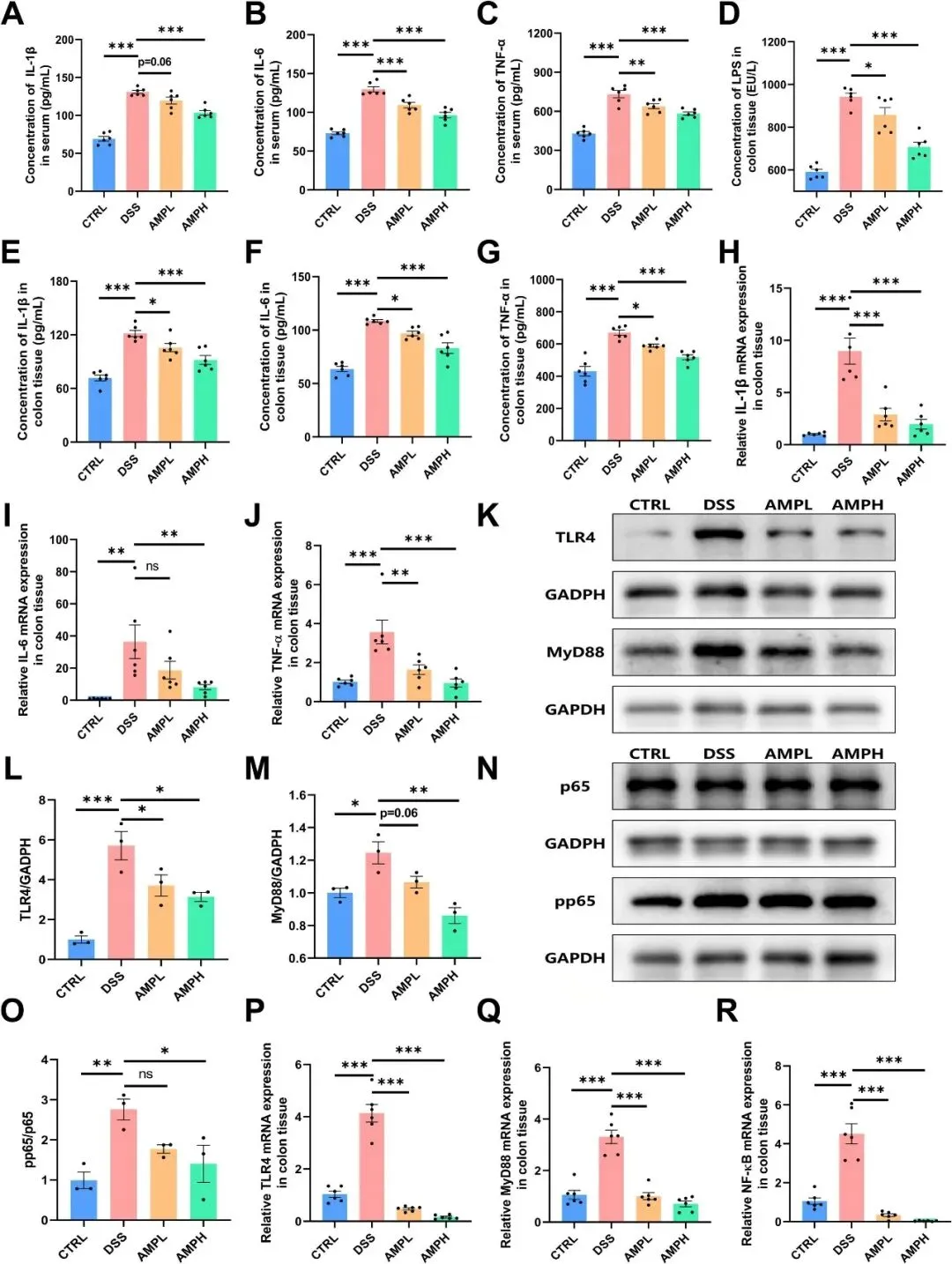
图5 AMP对细胞因子的调节和TLR4/NF-κB通路。(A-C)IL-1β、IL-6、TNF-α的血清浓度;n=6。(D-G)结肠组织中LPS(D)、IL-1β(E)、IL-6(F)和TNF-α(G)的浓度;n=6。(H-J)结肠组织IL-1β(H)、IL-6(I)和TNF-α(J)的mRNA表达;n=6。(K-O)结肠组织中TLR4、MyD88、p65和pp65的代表性蛋白质印迹带和密度分析结果;n=3。结肠组织中TLR4(P)、MyD88(Q)、NF-κB(R)的mRNA水平;n=6。
6. AMP改善DSS诱导的小鼠肠道功能障碍
DSS引发的肠道失衡在小鼠结肠炎的进展中起着核心作用。因此,研究者通过16S rRNA基因测序实现了结肠样本中的微生物分析。alpha多样性分析(图6A)显示,AMP增强了微生物的丰富度,但没有显著影响整体群落多样性。对于β多样性,NMDS分析显示CTRL、DSS和AMP干预组之间存在明显的分离,而AMPL和AMPH组显示出部分重叠,表明AMP干预部分恢复了DSS破坏的微生物组成(图6B)。门系水平分析表明,DSS降低了厚壁菌门,增加了拟杆菌门,降低了厚壁菌门/拟杆菌门(F/B)比率,而AMP给药部分恢复了这些变化(图6C-D)。在属水平上的进一步分析,结合LDA,确定了6个关键属(图6E-G)。具体而言,DSS增加了拟杆菌属、Escherichia-Shigella、副拟杆菌属和Clostridia vadinBB60簇的丰度,AMP可剂量依赖性地降低这些丰度。同时,AMP对Ruminococcus和Christensenellaceae R-7group的丰度有正向调节作用。总的来说,这些发现表明AMP可以选择性地调节特定的细菌属,以缓解DSS诱导的肠道生态失调。
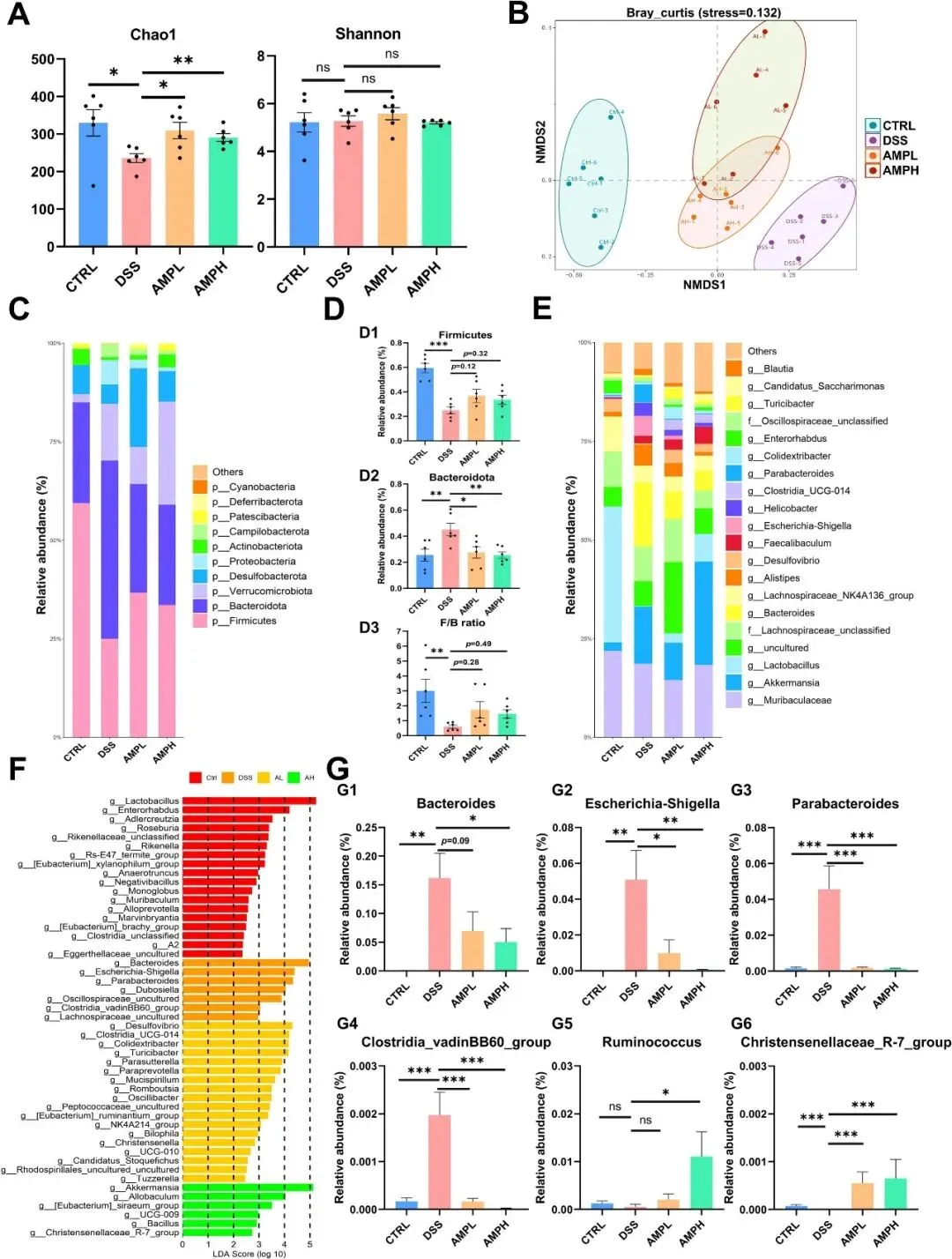
图6 AMP改善DSS诱导的小鼠肠道生态失调。(A)α-多样性度量(Chao1,Shannon)。(B)β多样性的NMDS图(Bray-curtis距离矩阵)。(C)前10个门水平微生物群落组成。(D)厚壁菌门和拟杆菌门的水平,以及F/B比值。(E)前20名微生物属水平组成。(F)LEfSe图(LDA>2.0)。(G)优势属比例。G1:拟杆菌,G2:Escherichia-Shigella菌,G3:副拟杆菌,G4:Clostridia vadinBB60簇,G5:Ruminococcus,G6:Christensenellaceae R-7group;n=6。
7. AMP改善DSS诱导的小鼠UC症状取决于肠道微生物群
在UC进展过程中,小鼠肠道微生物群落发生了变化,破坏了微生物稳态,进一步加剧了UC的进展。因此,研究者无法确定AMP是否直接通过微生物群调节直接发挥保护作用。研究者进一步进行了FMT实验,以了解AMP调节的小鼠肠道微生物群的影响。为了确保微生物群移植的稳定性,所有组的所有小鼠在实验前接受了为期7天的ABX治疗。具体实验步骤如图7A所示。研究结果表明,AMPH组的粪便微生物群显著改善了DSS模型小鼠的结肠缩短和脾脏肿胀(图7C-F)。H&E结果显示,AMPH+FMT组显著减轻了DSS诱导的肠组织损伤(图7H-I)。AMPH+FMT组没有改善DSS诱导的小鼠的体重变化,这与之前实验中AMPH组的结果不同(图7B)。总之,从AMPH组小鼠移植肠道微生物群可以缓解小鼠UC的发生。总的来说,这些数据表明,AMP在UC模型小鼠中的有益功效主要取决于AMP调节的肠道微生物群。
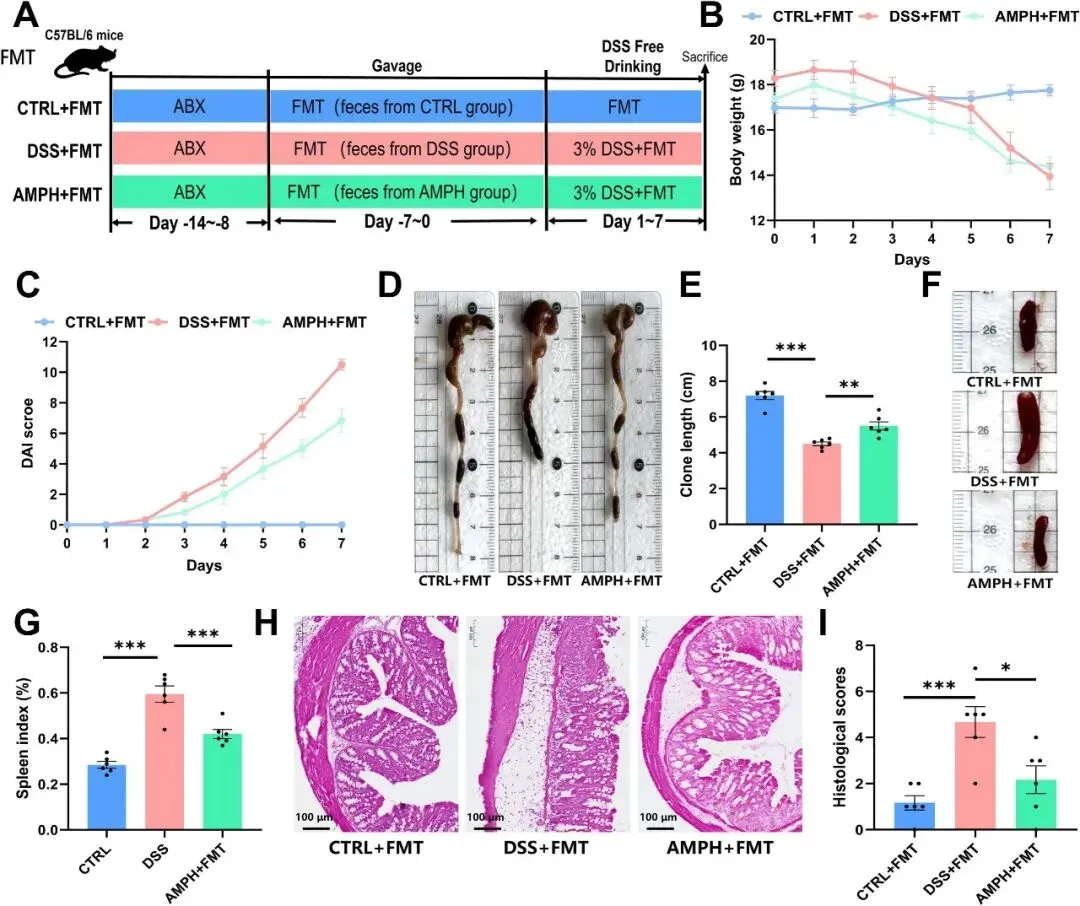
图7 FMT对小鼠UC的影响。(A)FMT实验示意图。(B)体重变化曲线,(C)DAI评分,(D)结肠取样形态,(E)结肠长度,(F)脾脏取样形态,以及(G)脾脏指数;n=6。(H)每组小鼠结肠的H&E染色图像。(I)组织学评分;n=6。
8. AMP可被肠道微生物群利用以产生SCFA
在确认AMP的功能依赖于肠道微生物群后,研究者进一步研究了肠道微生物群的相关代谢产物。多糖主要被肠道细菌分解成SCFA。因此,研究者使用气相色谱法测量了结肠内容物中的SCFA水平。如图8A所示,DSS建模显著降低了SCFA水平,而AMP抵消了这一影响。AMP治疗显著增加了结肠物质中乙酸、丙酸、丁酸、异丁酸和异戊酸的浓度(图8B-I)。其中,AMPH治疗组乙酸含量为DSS组的2.37倍,丙酸为2.58倍,丁酸为1.37倍,且丙酸含量略高于CTRL组。特征菌群与SCFA的相关性分析显示,拟杆菌属、志贺氏菌属、副拟杆菌属和Clostridia vadinBB60组与SCFA水平呈负相关,而Ruminococcus和Christensenellaceae R-7组与丙酸呈正相关(图8J)。为了进一步确认AMP是否可以转化为SCFA,研究者收集了小鼠粪便进行体外发酵实验。发酵48小时后的结果表明,AMP可以被细菌群落转化为SCFA(图8K-R)。与菊粉相比,AMP发酵产生的SCFA含量更高(不包括乙酸)。这些结果表明,肠道微生物将AMP转化为可观数量的SCFA,并有效地恢复结肠内容物中的SCFA水平。
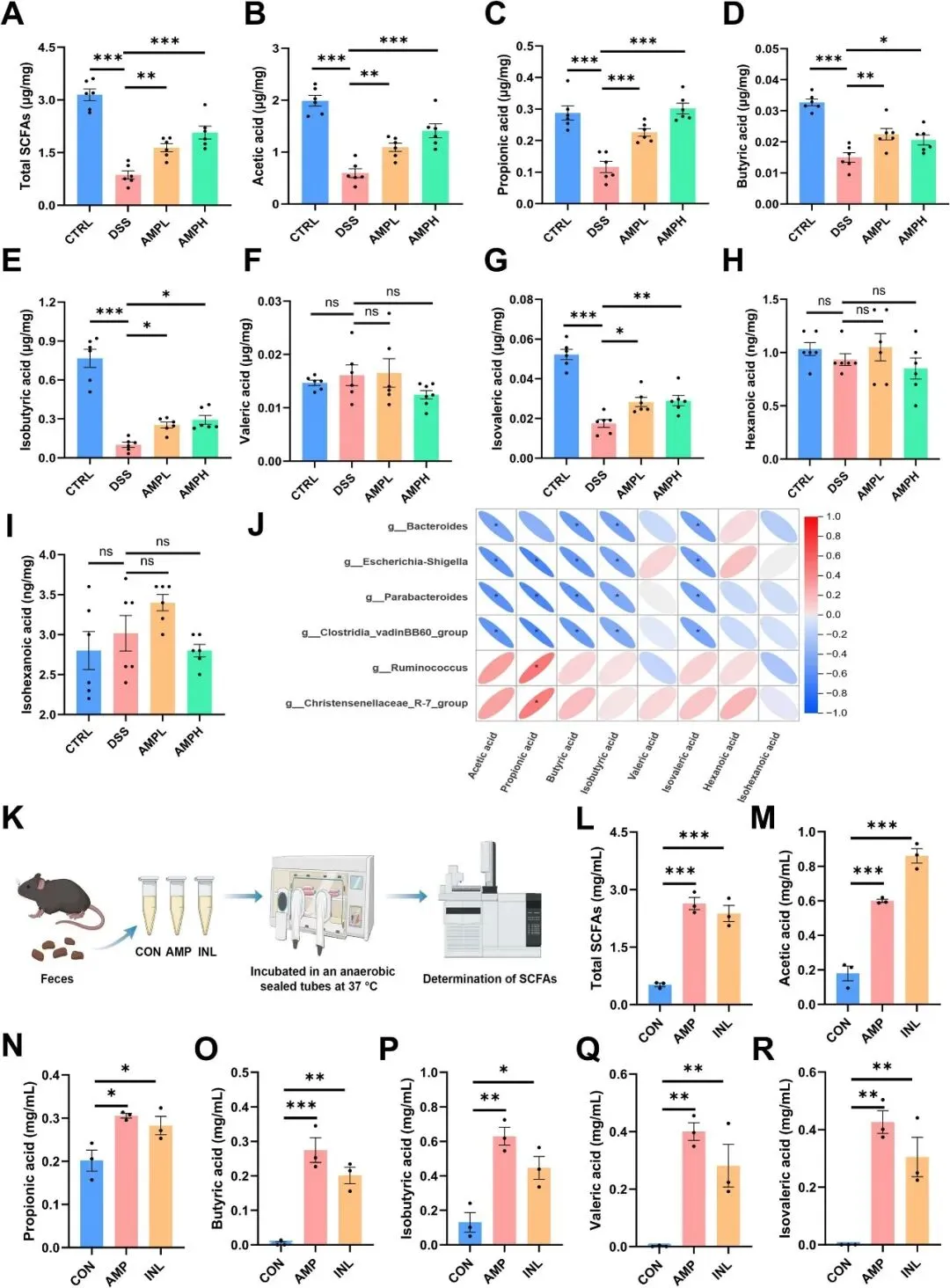
图8AMP可增加体内和体外的SCFA水平。(A)4组结肠内容物中的SCFA水平,n=6。(B-J)乙酸(B)、丙酸(C)、丁酸(D)、异丁酸(E)、戊酸(F)、异戊酸(G)、己酸(H)和异己酸(I)的浓度。(J)SCFA微生物组生物标志物相关性热图。(K)体外发酵过程示意图。(L)发酵48小时后的总SCFA水平,n=3。(M-R)发酵48小时后乙酸(M)、丙酸(N)、丁酸(O)、异丁酸(P)、戊酸(Q)、异戊酸(R)的浓度,N=3。
9. AMP改善DSS诱导的小鼠结肠组织代谢紊乱
作为关键的肠道微生物群副产物,SCFA显著影响宿主代谢调节。DSS建模诱导SCFA相关受体GPR41、GPR43和PPARγ的转录下调,AMP逆转了这些变化(图S9)。为了研究AMP的代谢作用,研究者对小鼠结肠组织进行了代谢组学分析。PLS-DA在DSS队列和各种其他实验亚群中显示出代谢物模式的显著差异,而AMPL和AMPH组显示出高度相似的代谢物组成(图9A)。研究者使用阈值为VIP>1.0、FC>1.5和P<0.05的火山图,发现与DSS对照组相比,AMPH治疗小鼠中有21种代谢物上调,34种代谢物下调(图9B-D)。随后,通过京都基因和基因组百科全书(KEGG)途径富集对这些差异代谢物进行了注释和分析,证明AMP抵消了DSS诱导的几种代谢途径的干扰,如甘油磷脂、花生四烯酸、亚油酸代谢和初级胆汁酸生物合成(图9E-F)。详细信息如表4所示,每种差异代谢物的相对丰度如图S2所示。特征菌群与代谢产物的相关性分析显示,Escherichia-Shigella,Parabacteroides,和Clostridia vadinBB60组与LysoPC(15:0/0:0)、LysoPC和24-Hydroxycalcitriol呈正相关。而Ruminococcus和Christensenellaceae R-7group与腺苷和Taurocholicacid呈正相关,与LysoPC(15:0/0:0)、LysoPC(18:1/0:0)和花生四烯酸呈负相关(图S3)。总之,这些结果表明,AMP可能通过调节肠道微生物群组成来提高SCFA水平,最终缓解DSS诱导的肠道代谢紊乱。
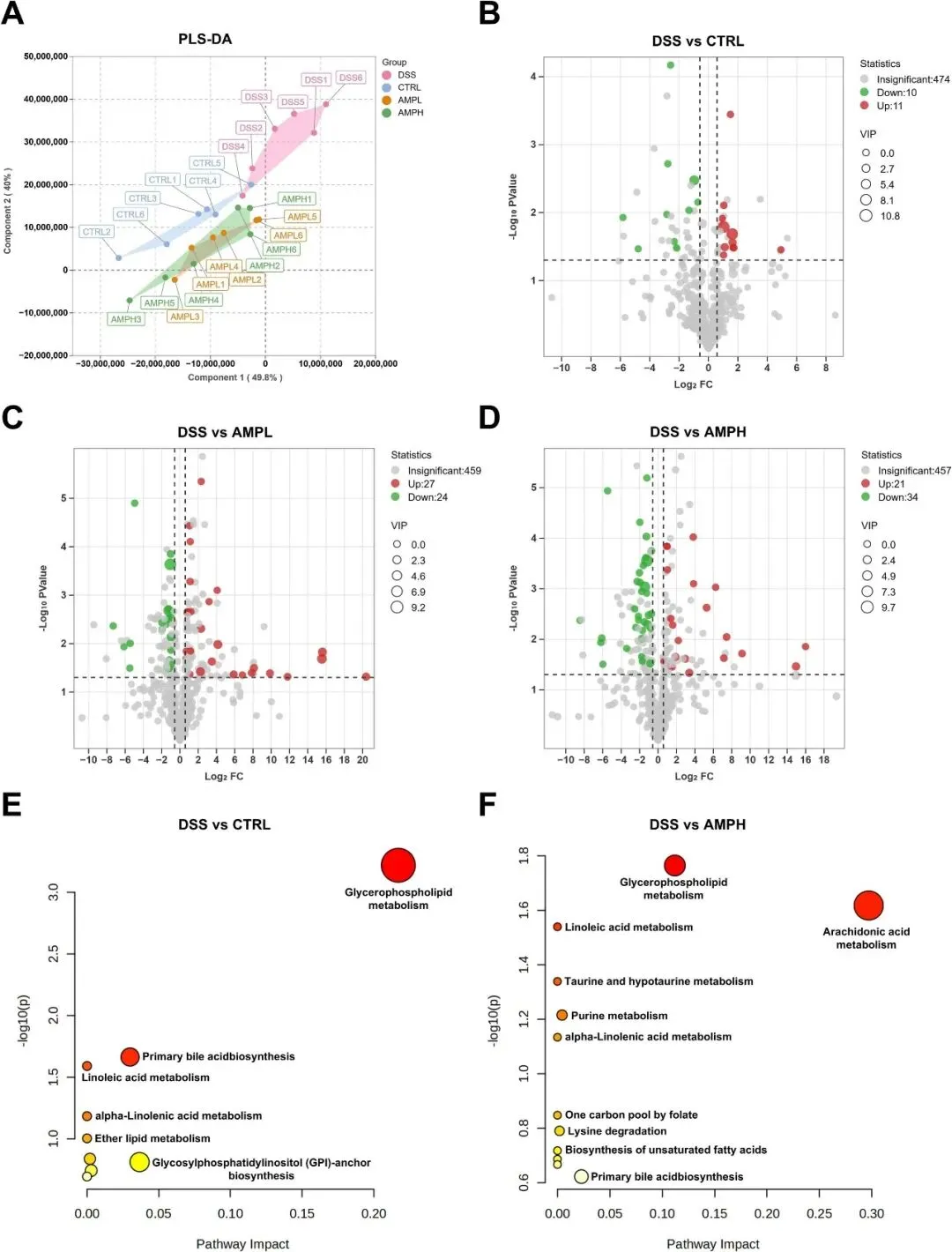
图9 AMP改善DSS诱导的小鼠结肠组织代谢紊乱。(A)PLS-DA图显示了4组结肠样本。(B-D)火山图(B:DSS vs CTRL;C:DSS vs AMPL;D:DSS vs AMPH)。(E-F)通路分析总结(E:DSS vs CTRL;F:DSS vs AMPH)。
表4 AMP对KEGG通路的影响。
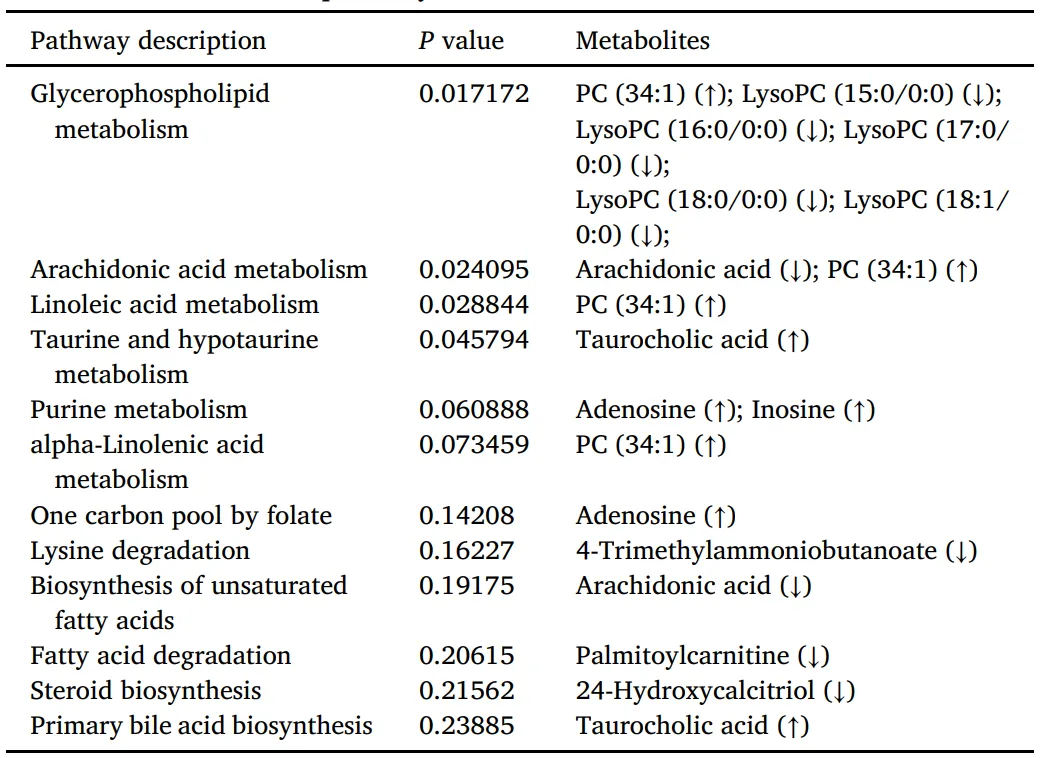
注:向上箭头(↑)表示AMPH组的代谢物相对丰度高于DSS组,而向下箭头(↓)表示丰度较低。
讨论
本项研究产生了AMP,一种来自AMB干燥根茎的纯化多糖,与之前报道的AMB多糖在结构上存在显著差异。Guo等人分离出五种由甘露糖和葡萄糖组成的纯化多糖,而Zhang等人报道了含有葡萄糖和阿拉伯糖的中性多糖,以及含有葡萄糖和半乳糖的酸性多糖。本研究阐明,AMP是一种中性多糖,主要含有果糖(95.99%)和少量葡萄糖。Qin等人报道了一种来自AMB的果聚糖,其具有→ 6)-β-D-Fruf-(2 →骨架,末端→1,6)-β-D-Fruf-(2 → 1)β-D-Fruf-(2 → 和O-6连接的侧链β-D-Fruf-(2 → 。相比之下,AMP具有以下核心链→1)-β-D-Fruf-(2 →和→1,6)-β-D-Fruf-(2 →连接,带有类似高度分支果聚糖的短β-D-Fruf-(2 → 6)-β-D-Fruf-(2 → 侧链。多糖的生物活性受到其结构特征的显著影响,包括单糖组成、糖苷键和分支模式。例如,葡萄糖含量影响益生元活性——果糖残基中C2异头碳的β构型增强了肠道消化的抵抗力,促进了肠道微生物群的利用;阿拉伯糖和木糖主要影响免疫调节活性。AMP由一种中性单糖组成,只含有羟基(-OH),没有酸性修饰,这证实了它是一种中性多糖。来自同一植物的中性和酸性多糖在功能上不同:酸性多糖具有成膜、水凝胶、控释和稳定特性,有利于药物递送系统的开发,而中性多糖被广泛报道具有抗炎、抗氧化和调节肠道微生物群的活性。
与线性结构相比,更高的支化度使多糖能够形成更多的功能结构域。例如,随着中性侧链数量的增加,果胶多糖对DSS诱导的结肠炎表现出更强的治疗效果。此外,支链多糖的免疫刺激活性明显高于线性多糖。研究表明,分支结构可以直接刺激巨噬细胞,表明分支本身可能充当免疫系统识别的模式分子。研究表明,高支链结构是其抗衰老活性的必要因素;这可能是由于低支链多糖不能提供足够的免疫识别位点。关于黄芪多糖的研究表明,增加分支可以提高体外抗肿瘤活性和免疫调节功能。此外,具有支链结构的果聚糖存在更多暴露的羟基,从而增加了它们在清除活性氧物种时提供氢或电子的潜力。羟基含量的增加同时有助于提高多糖的溶解度;相关研究表明,提高溶解度有助于肠道微生物群的发酵性能。因此,分支结构的存在可能会改变肠道微生物对果聚糖的降解模式和速率。此外,与直链果聚糖相比,支链果聚糖对乳杆菌等益生菌具有更强的促生长作用。因此,与线性果聚糖相比,AMP可能具有更好的抗炎和肠道微生物群调节功能。
多项研究结果表明,中草药多糖通过改善肠道保护机制改善UC的疗效。肠道屏障对维持肠道稳态至关重要,与各种炎症性疾病密切相关,包括感染、IBD和乳糜泻。它包括机械、化学、免疫和微生物屏障。由紧密连接、粘液和细胞连接组成的上皮屏障对于调节肠道通透性至关重要。紧密连接蛋白的遗传缺失会破坏肠道屏障并诱导UC样表型。正常情况下,杯状细胞分泌粘蛋白形成保护性粘液屏障;然而,结肠炎通常会损害这种粘液屏障。据报道,人参和黄蜀葵中的多糖可以上调紧密连接蛋白或MUC2的表达,从而修复屏障损伤并缓解结肠炎。研究结果表明,AMP显著提高了小鼠结肠组织中紧密连接蛋白的转录和表达水平。DSS主要通过破坏肠道屏障诱导结肠炎,使LPS穿透体循环,激活TLR4/NF-κB通路,引发组织炎症。TLR4特异性识别入侵细菌的LPS,招募衔接蛋白MyD88,并激活NF-κB,NF-κB是参与炎症和屏障调节的关键转录因子。过度的NF-κB激活会损害肠道屏障,加剧UC的进展。总的来说,本研究结果表明,AMP通过增强肠屏障的完整性和抑制TLR4/NF-κB信号通路来减轻组织炎症。
作为微生物屏障的组成部分,肠道微生物群通过调节免疫反应来支持肠道稳态和整体健康。肠道微生物群失衡或生态失调会破坏肠道稳态,损害肠道屏障,引发局部和全身炎症,削弱免疫力。在无菌条件下饲养的IL-10-/-缺陷小鼠不会患结肠炎,而暴露于传统微生物的小鼠会患结肠炎,这突显了肠道细菌在引发炎症中的关键作用。调整肠道微生物群组成具有越来越大的潜力,并被认为是UC管理的一大希望。举例来说,人参皂苷化合物K改善了啮齿动物的UC,这一益处与Akkermansia、Candidatus Saccharomonas和Ruminococacae UCG-014分类群的增加有关。同样,中草药多糖可以通过恢复肠道免疫平衡和调节肠道微生物群落对UC产生药理作用。FMT实验表明,AMP的抗结肠炎功效取决于肠道微生物群。在门阶段,F/B比例的变化是生态失调的标志,通常在IBD病例中具有较低的比例。与这些报告一致,本研究中的DSS降低了F/B比值,而AMP干预逆转了这一效果。在属水平上,DSS提高了拟杆菌属、Escherichia-Shigella、拟副杆菌属和Clostridia vadinBB60组的数量,而AMP剂量降低了这些数量。拟杆菌属是拟杆菌科的一个主要属,可以携带抗生素抗性基因,引发持续的粘膜免疫反应,通过外源性益生菌使UC治疗复杂化。AMP可以通过调节内源性微生物群来减轻这种风险。Escherichia-Shigella是一种常见的病原体,其在UC患者中的扩散通过LPS释放促进肠道炎症。副拟杆菌具有促炎作用和潜在的肠道致病性。相反,AMP对Ruminococcus和Christensenellaceae R-7组的丰度有正向调节作用。食用富含纤维,特别是抗性淀粉的饮食会促进Ruminococcus的增殖,Ruminococcs是SCFA产生的关键因素,尤其是丁酸盐。Christensenellaceae R-7组在IBD患者中显著减少,有助于肠道微生物群的稳定,并参与碳水化合物发酵,产生乙酸和丁酸。AMPH+FMT组观察到的体重减轻可能是由于AMP本身作为发酵底物,提供额外的营养支持,有助于维持肠道代谢和微生物稳态。这些结果共同表明,AMP通过重建肠道微生物群平衡来影响抗溃疡性结肠炎作用。
肠道微生物群产生了一系列对人类发展和整体健康至关重要的代谢物。研究表明,果聚糖是微生物发酵的关键底物,产生大量的SCFA,同时促进有利细菌菌株的增殖。本研究观察到,在AMP给药后,UC小鼠结肠内容物中的SCFA增加。其中,AMP主要代谢为乙酸和丙酸,其次是丁酸,这与主要产生丁酸的果聚糖的常见体内发酵模式不同。研究结果表明,AMP治疗可以减少DSS引起的代谢紊乱,包括甘油磷脂、花生四烯酸和亚油酸代谢紊乱。在UC进展过程中,甘油磷脂代谢和亚油酸代谢的紊乱会激活炎性细胞释放细胞因子,进一步损害肠道屏障功能并加剧炎症。甘油磷脂代谢可以增加甘油磷脂向花生四烯酸的转化,导致过度积累、NF-κB通路激活和肠道屏障损伤。SCFA可以通过GPR43受体信号激活PPARγ,从而调节脂肪酸摄取、酯化和磷脂合成,调节甘油磷脂和花生四烯酸代谢,减轻炎症。SCFA还可以通过调节TLR4/NF-κB通路来缓解肠道炎症。Pan等人证明,醋酸盐通过GPR41和GPR43发挥作用,维持结肠炎小鼠杯状细胞数量和肠屏障完整性,同时通过上调GPR41抑制NF-κB活化。Zhang等人发现,醋酸盐和丙酸盐还可以通过直接结合MyD88并降低肠道中TLR4、MyD88和NF-κB p65的表达水平来缓解肠道炎症。Hazime等人证实,丁酸盐可以逆转肠道屏障蛋白的下调,减少肠道炎症,纠正生态失调。此外,PPARγ可以被丁酸盐直接激活,PPARβ的激活可以促进脂肪酸氧化,抑制NF-κB活性,维持上皮屏障功能。特征性肠道微生物群、SCFA和代谢产物之间的相关性分析表明,AMP可能通过特征性调节肠道微生物群并转化为SCFA来发挥其抗结肠炎作用,从而改善肠道代谢紊乱,恢复肠道屏障,抑制TLR4/NF-κB信号通路的激活。
未来的研究将集中在AMP衍生的功能性产品的开发上,包括结构修饰和工艺优化。此外,本研究的目标是整合宏基因组测序、靶向代谢组学和空间代谢组学,以鉴定特征微生物类群和由肠道微生物群调节的特定差异代谢物。这种方法将使研究者能够更深入地研究AMP治疗作用的确切机制。
结论
在本研究中,研究者从薤白中提取了一种以前未鉴定的多糖(AMP)。结构分析表明,AMP主要由果糖和葡萄糖组成,具有复杂的分支结构。主干由以下部分组成→1)-β-D-果糖-(2→和→1,6) -β-D-Fruf-(2→连接。在DSS治疗的结肠炎小鼠中,AMP通过抑制潜在致病菌属和促进SCFA产生细菌来恢复肠道微生物平衡并增加SCFA浓度。此外,AMP调节结肠上皮细胞中的关键代谢过程,介导SCFA相关的保护机制。此外,急性毒性试验表明,AMP具有较大的安全度。本研究的数据揭示了AMP作为功能性成分治疗溃疡性结肠炎的前景,证实了推进AMP衍生功能性产品的科学依据。
----------微科盟精彩文章----------
江苏省中医沈洪发文揭示白芷多糖通过降低肠道菌源咪唑丙酸从而激活PPAR-γ信号改善结肠炎
Int J Biol Macromol :山药多糖通过调节肠道微生态和代谢紊乱改善溃疡性结肠炎
如果需要原文pdf,请扫描文末二维码



获取此文献原文PDF、申请加入学术群,联系您所添加的任一微科盟组学老师即可,如未添加过微科盟组学老师,请联系微生态老师9,无需重复添加。
请关注下方名片,了解更多代谢组学知识

