免疫哨兵的“意外”代价
故事始于一个名为CARD9的免疫蛋白。作为机体识别微生物的关键接头蛋白,CARD9是维持肠道真菌平衡的重要哨兵。研究团队观察到,当小鼠缺失CARD9基因后,即便同样进食高脂饮食,它们也表现出更严重的代谢紊乱。与正常小鼠相比,这些基因敲除鼠体重显著增加,肝脏甘油三酯含量飙升,脂肪肝评分恶化,且胰岛素敏感性严重受损。
转录组分析直指问题的核心:脂肪酸氧化通路失灵。在CARD9缺失小鼠的肝脏中,负责燃脂的关键基因和蛋白,如CPT1A和ACOX1,其表达被显著抑制。这相当于细胞的“能量工厂”线粒体减慢了运转,导致脂肪无法被有效消耗,大量堆积在肝细胞内。
“嫌疑人”并非细菌
为了揪出幕后推手,团队设计了一系列精巧的验证实验。他们首先对小鼠使用了广谱抗生素和抗真菌药物,再利用粪菌移植技术进行回补。结果显示,只有当肠道真菌被清除后,CARD9缺失带来的脂肪肝加剧现象才完全消失。而单独清除细菌,却未能逆转这种表型差异。
这一关键证据将矛头指向了肠道真菌群落的失衡。通过对粪便的ITS测序,一种名为酿酒酵母的真菌浮出水面。在CARD9缺失小鼠的肠道中,酿酒酵母的相对丰度出现了特异性锐减。团队推测,这种常见于面包和啤酒发酵的微生物,很可能就是那个缺失的保护因子。
一条跨越肠肝的信号通路
验证推测的方式直接而有效。研究团队每隔三天给高脂饮食小鼠灌胃酿酒酵母。结果显示,补充酿酒酵母即便未能让其在肠道内永久定植,也足以产生显著的保护效果。这不仅阻止了小鼠体重飙升,减轻了肝脏脂肪变性,还将受损的脂肪酸氧化效率重新激活。
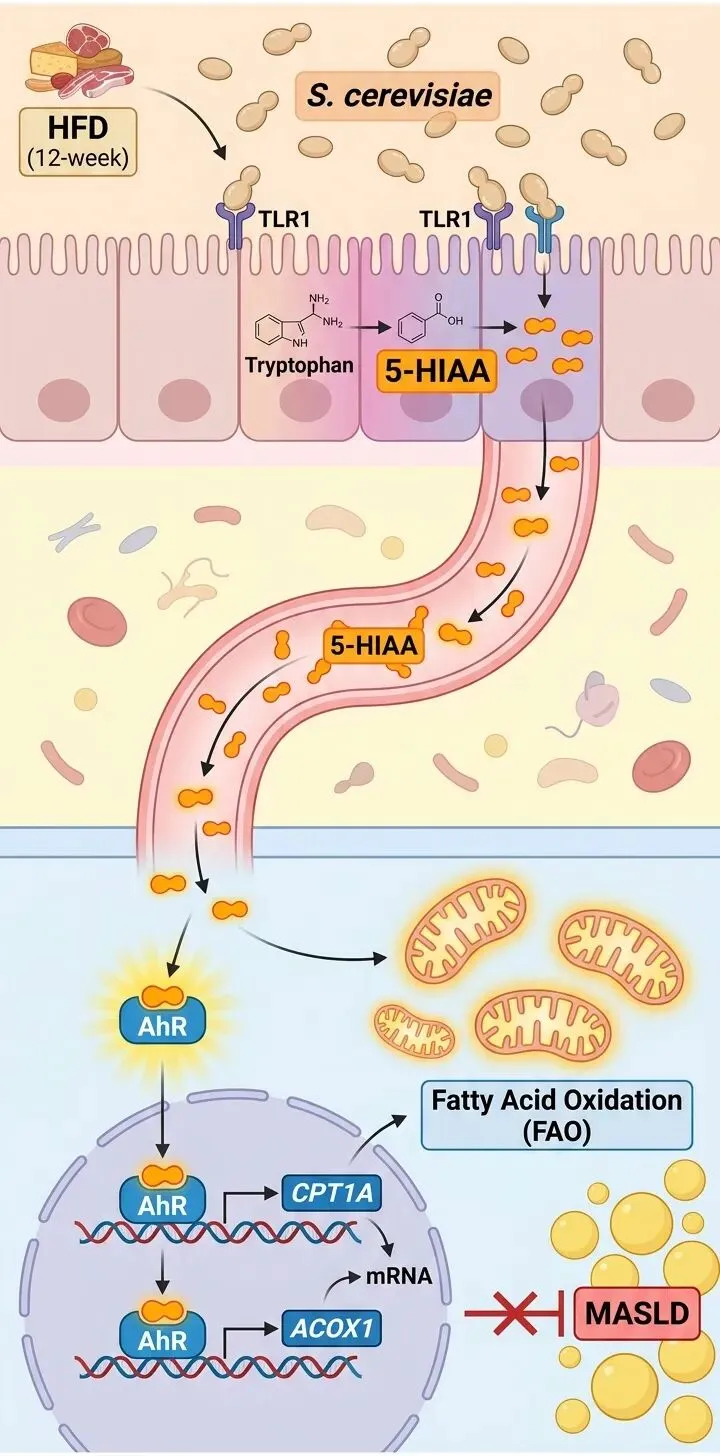
更有趣的是,热灭活的酿酒酵母同样保留了类似的功效。这让研究者意识到,发挥作用的很可能不是活菌本身,而是其细胞壁上的某些热稳定性成分。机制探究由此指向了模式识别受体。他们发现,酿酒酵母通过激活肠道上皮细胞的TLR1受体,精准调控了色氨酸向5-HIAA的代谢流向。
5-HIAA的“燃脂指令”
5-HIAA是色氨酸在血清素通路中的下游产物。尽管酿酒酵母自身并不直接产生5-HIAA,但它能通过TLR1-NF-κB信号轴,持续上调肠道细胞中单胺氧化酶的表达,从而源源不断地将血清素转化为5-HIAA。这些代谢物随后经门静脉入肝,发挥核心调控作用。
在肝细胞层面,5-HIAA扮演了“燃脂开关”的角色。研究证实,5-HIAA能够作为配体直接结合芳香烃受体。结合后的AhR迅速向细胞核内转移,随后结合到CPT1A和ACOX1基因的启动子区域,精确启动这些基因的转录。这如同给肝细胞下达了明确的燃脂指令,线粒体的最大呼吸能力和备用呼吸能力随之大幅增强。
从临床验证到未来挑战
基础研究的成果能否经受临床检验至关重要。研究团队一方面对临床上收集的MASLD患者样本进行分析,确认了患者肠道中酿酒酵母丰度和体内5-HIAA水平均显著低于健康人群,且5-HIAA浓度与肝损伤指标呈现负相关。
另一方面,他们在一项小规模的双盲对照临床试验中,让MASLD患者连续8周补充酿酒酵母。结果显示,治疗组的空腹血糖、胆固醇和转氨酶水平均出现下降。更为直观的是,通过超声衰减分析和磁共振质子密度脂肪分数扫描,患者的肝脏脂肪含量切实得到了改善。
尽管前景光明,研究者也审慎指出了现有工作的局限。例如,全身性的CARD9敲除可能伴随复杂的免疫背景,与体外癌细胞系模型一样,需要未来在组织特异性敲除动物和更接近生理状态的原代模型中进一步确证。此外,5-HIAA除了激活AhR,还可能作用于GPR35等其他受体,其下游机制网络仍有待完整描绘。
不过,这项研究无疑为“肠-肝轴”的概念注入了崭新的内涵。它让学界认识到,在以细菌为主角的肠道微生态舞台上,真菌同样举足轻重。未来,锁定酿酒酵母中的活性分子作为后生元,或者单独补充5-HIAA类似物,极有可能为全球日益严峻的脂肪肝流行提供一种安全且精准的干预良方。