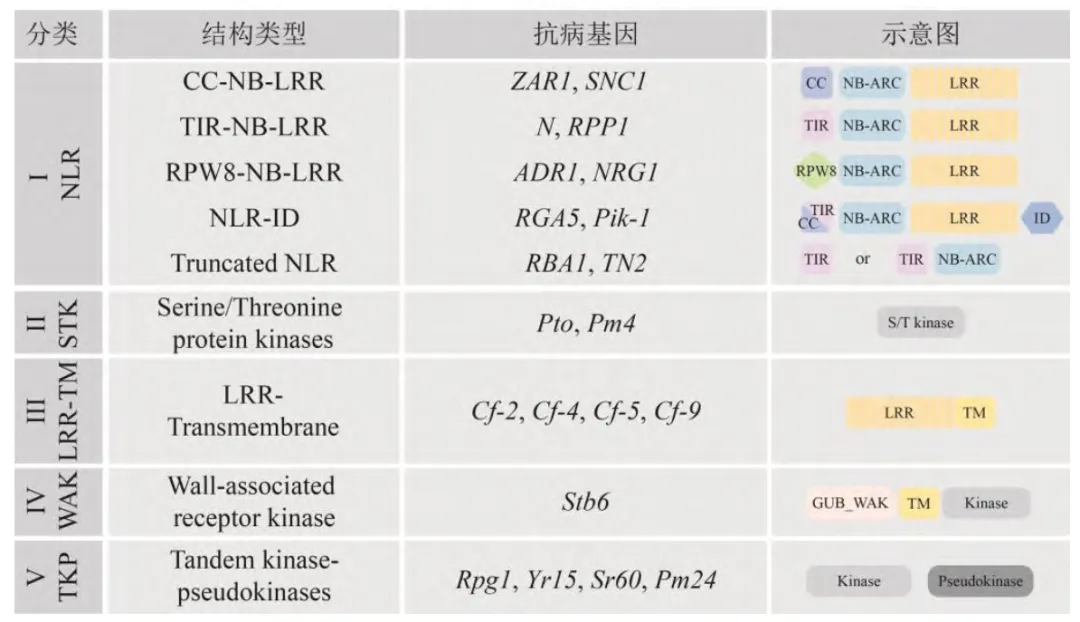
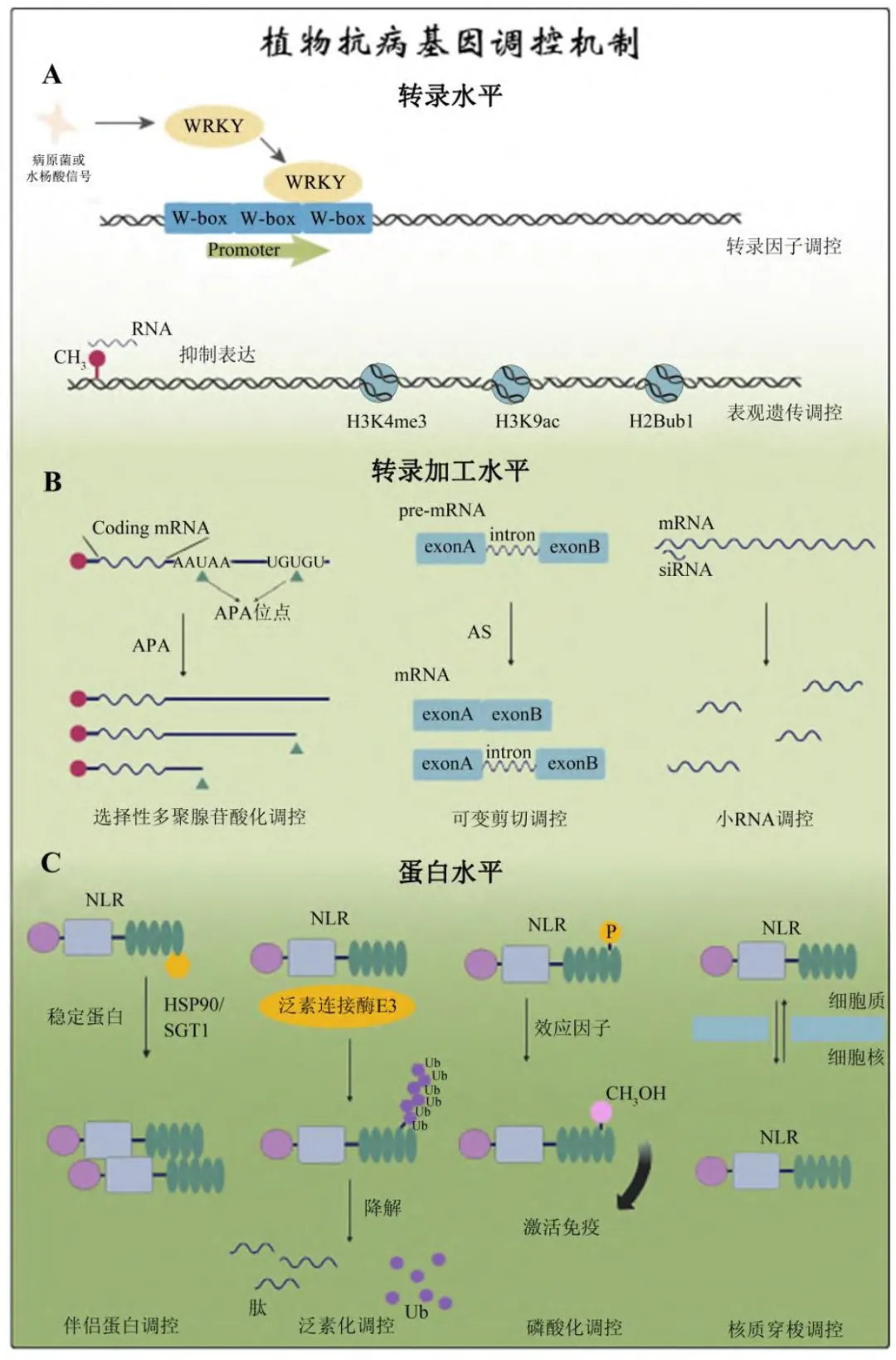
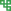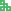抗病基因的组织特异性及时空特异性表达与其功能紧密相关[37]。转录水平上的调控主要包括转录因子(transcription factors)以及与转录因子结合的顺式作用元件(cis-acting element)。除此之外,包括DNA修饰(DNA甲基化)、组蛋白修饰(组蛋白甲基化、乙酰化、泛素化) 以及染色质重塑在内的表观遗传修饰也在抗病基因的转录调控上发挥着重要作用。
1.1 转录因子调控
转录因子通过与顺式作用元件特异性结合调节基因转录活性, 在植物响应病原菌侵染过程中起着至关重要的作用。WRKY家族是植物最大的转录因子家族之一,能够特异性结合下游基因启动子W-box顺式作用元件调控基因转录水平,在植物免疫中发挥着重要作用[38]。Mohr等人发现W-box元件在植物抗病基因启动子序列中显著富集,其中拟南芥抗病基因RPP8的启动子区域存在三个W-box基序,并且该基因的表达受到病原菌及水杨酸的诱导。当突变三个 W-box调控功能域后,抗病基因RPP8的表达水平无法被病原菌诱导上调, 导致植物抗病性下降,表明抗病基因RPP8启动子区W-box基序对于其表达水平及抗性激活具有重要作用,同时W-box在抗病基因中的保守存在暗示了WRKY转录因子对于抗病基因表达以及免疫调控的潜在意义[39]。锌指类转录因子在抗病基因转录调控中也发挥着重要作用。水稻锌指蛋白 OsVOZ1和OsVOZ2可结合并促进抗病基因Piz-t的表达水平, 以提高 Piz-t 介导的稻瘟病抗性[40]。拟南芥抗病基因SNC1的精细表达对生长和免疫的平衡具有重要意义,通过筛选 snc1突变体的抑制子获得了多个mos(modifier of snc1)突变体,发现 MOS1通过与转录因子TCP15结合增强其与SNC1启动子的结合进而促进 SNC1的表达。除转录因子之外, 多个位于启动子区和下游非编码区的顺式作用元件也共同调控SNC1的表达水平[41, 42]。由于抗病基因的转录对激活抗病反应至关重要, 因而在与病原菌的军备竞赛中,植物抗病基因进化出一种“诱饵型”顺式作用元件,用于快速响应病原菌侵染。TALE(TAL effector)是一类最初从黄单胞菌(Xanthomonas spp.) 鉴定到的转录因子类效应分子, 通过结合并促进植物感病基因的表达, 以提高病原菌的致病力。相应地,植物抗病基因启动子区域进化出一类特定基序,作为“诱饵”被TALE结合并促进抗病基因表达。例如,辣椒抗病基因Bs3启动子区域进化出黄单胞菌TAL 效应分子 AvrBs3 的结合基序, 黄单胞菌分泌 AvrBs3 进入植物细胞, 结合并激活Bs3的表达, 从而引发植物抗病反应[32]。目前, 已报道多个抗病基因的表达和激活受到病原菌分泌的TAL效应分子的转录调控, 如大麦抗病基因Rph3[31],水稻白叶枯抗病基因 Xa10、 Xa23、 Xa27[43-45], 以及辣椒抗病基因Bs4C等[46]。
1.2 表观遗传调控
DNA甲基化是表观遗传水平上的修饰之一,这一修饰存在于所有高等生物中,并与基因表达调控密切相关。研究发现植物抗病基因广泛存在DNA甲基化修饰, 并且该修饰参与调控植物抗病反应[47]。全基因组甲基化水平分析发现菜豆NLR抗病基因中约一半存在甲基化修饰, 远高于其他基因家族。其中,约有 90个被24 nt的siRNA靶向, 且这些NLR基因的表达水平很低甚至不表达,表明其受RdDM(RNA介导的DNA甲基化)介导的基因沉默调控[48]。除全基因组水平外,一些抗病基因的表达调控也与DNA甲基化修饰高度相关。水稻抗病基因PigmR能够对稻瘟病不同生理小种提供广谱抗性, 但过量表达或持续激活严重影响水稻生长与产量,同一位点的PigmS能够与与 PigmR 互作形成异源二聚体并抑制 PigmR 介导的抗病反应。研究发现PigmS的组织特异性表达受 RNA介导的DNA甲基化调控, 叶片中PigmS启动子区域存在较高的甲基化水平并且表达量较低,而在花粉中该基因发生甲基化水平较低因此表达量水平较高,通过调控不同组织中Pigm的表达水平实现水稻产量和抗病之间的平衡[10]。水稻抗病基因Xa21G启动子区域存在5mC甲基化修饰导致其无法表达, 而在人工去甲基化处理的突变体中,Xa21G启动子区域的甲基化被消除恢复其自身表达量,表现出对黄单胞菌水稻致病变种的抗性[49]。有研究发现5mC甲基化修饰对抗病基因的表达也具有调控功能,研究发现水稻抗稻瘟病基因Pib启动子区域 5mC甲基化修饰与Pib的表达水平呈正相关[50]。因此, 不同类型的甲基化修饰对抗病基因的表达及功能具有重要的调控作用,高水平的甲基化往往能够降低抗病基因的转录水平,减少细胞的能量消耗,从而维持正常的生长。相反, 当病原菌攻击时抗病基因也能通过甲基化调控增加自身表达量, 快速激活植物对病原菌的抗性[51]。组蛋白作为核小体的基本组成成分能够发生多种化学修饰,如甲基化、乙酰化、泛素化等。这些修饰通过调控染色质的状态进而调控基因的表达及稳定性,在植物的生长发育及外界信号响应过程具有重要作用[52]。例如,H3K4甲基转移酶ATXR7通过调控抗病基因SNC1以及RPP4基因上的H3K4me3水平增强抗病基因的表达量,进而激活拟南芥对霜霉病的抗性[53]。组蛋白甲基转移酶 SDG8参与调控拟南芥抗病基因LAZ5的H3K36me3修饰水平以持续激活LAZ5 转录活性,进而影响植物免疫[54-56]。高粱抗病基因ARG15末端非翻译区(5’UTR) 的H3K36me2及H3K36me3的水平在病原菌侵染过程中显著富集, 导致ARG表达水平上调从而激活高粱的抗性水平;而在高粱感病品种中, ARG1 基因上H3K36me2/me3 水平较低,导致基因表达水平较低,无法对病原菌提供完全抗性[57]。组蛋白乙酰化对NLR基因的表达也具有调控作用, 例如, 组蛋白去乙酰化酶(HDACs)HDA9与其互作蛋白HOS15能够调控多个拟南芥抗病基因的H3K9乙酰化修饰水平, 进而调控抗病基因表达以达到抗病与生长发育平衡[58]。除此之外,E3泛素连接酶HUB1和HUB2介导SNC1位点上的H2B泛素化,在病原菌侵染时促进SNC1的表达, 并与组蛋白甲基化和乙酰化共同调控SNC1的表达[59, 60]。
染色质重塑发生在基因的复制和重组过程, 是指染色质包装状态、 核小体中组蛋白以及对应DNA分子发生的改变。染色质重塑复合物由染色质重塑因子组成, 分为四大类:SWI/SNF、 CHD、 ISWI 和 INO80[61]。研究发现染色质重塑复合物对于抗病基因的表达具有重要调节作用。BAF60 是SWI/SNF染色质重塑复合物中的一个亚基, 可通过ATP水解产生的能量在DNA上定位核小体[62],Chien-Yu Huang等人发现BAF60在拟南芥中的同源蛋白SWP73A能够结合抗病基因RPS2启动子区域调控H3K9me2水平以抑制RPS2的表达。当病原菌侵染时,miR3440和siRNA-SWP73A靶向SWP73A导致该基因沉默,因此激活了RPS2的表达以及拟南芥的抗病反应[63]。Zou等人发现染色质重塑蛋白 CHR5作为CHD染色质重塑复合体亚基,可调控核小体占有率,参与调控 SNC1的表达水平[64]。此外,拟南芥SWI/SNF染色质重塑复合体SYD也在在染色质水平上抑制SNC1的转录 [65]。2 RNA 水平调控
基因的表达调控不仅发生在转录水平, 在转录后水平上也存在多种调控方式。转录后调控(post-transcriptional regulation)是指发生在RNA水平上的调控作用,具体包括pre-mRNA 的多聚腺苷酸化,剪切加工,mRNA 的稳定性调控, RNA干扰介导的转录后基因沉默等[66]。2.1 选择性多聚腺苷酸化调控
转录后mRNA 在polyA加尾时可能会选取不同的位置,使得一个基因可产生不 同3’UTR的转录本,也被称为选择性多聚腺苷酸化(Alternative polyadenylation,APA)。APA普遍存在于真核生物中,参与调控基因转录本的多样性和稳定性。研究发现,拟南芥开花相关基因 FLC、FPA 及FCA 的表达水平都受到APA的调控作用[67]。抗病基因表达模式分析发现拟南芥中72个抗病基因上都鉴定到APA,表明该调控在抗病基因上的普遍存在,但具体功能还不清楚[37]。2017 年,Tokuji Tsuchiya等人发现RPP7的5’UTR 区域插入了一个转座子该转座子插入位置携带 H3K9me2 修饰,甲基化修饰水平的变化导致 RPP7 的多聚腺苷酸化位点发生改变。遗传分析发现,EDM2蛋白参与了RPP7基因的选择性多聚腺苷酸化, 直接调控 RPP7 具有抗性功能的转录本的表达水平, 对其介导的抗病反应至关重要[68]。2.2 前体 RNA 加工调控RNA剪切是指前体 RNA(pre-mRNA) 中内含子去除, 外显子连接的过程,这一过程由剪切复合体完成。剪切复合体是一种进化上保守的核糖核蛋白复合物,通过识别内含子结构并对其剪切产生成熟的mRNA, 这一过程可能会发生可变剪切, 导致同一基因产生多种转录本。这种调控方式在植物中普遍存在, 已知拟南芥60%的非单个外显子基因均存在可变剪切调控[69],而其他双子叶及单子叶植物中也有 30%-50%的多外显子基因存在可变剪切调控[70-72]。
可变剪切对于蛋白多样性和丰富性具有重要意义[73], 大量研究发现植物抗病基因受到可变剪切调控。Mandadi等人利用RNA-seq 数据对二穗短柄草(Brachypodium distachyon) 在病毒侵染过程中可变剪切事件进行分析, 发现 100多个免疫相关基因在病毒侵染过程中发生可变剪切, 其中包含9个抗病基因[74]。Zhang 等人对拟南芥剪切因子SR45的功能进行分析,多个抗病基因在内的542个基因发生的可变剪切受到SR45的调控, 因此 sr45-1 突变体植物表现出增强的抗病表型可能与这些抗病基因的表达水平相关[75]。这些研究表明植物抗病基因可变剪切广泛存在, 并且对抗病基因功能的调控发挥重要作用。
烟草抗病毒基因N是植物中第一个被报道的存在可变剪切的抗病基因, 可变剪切产生的编码全长蛋白的转录本 NS 和编码截短蛋白的转录本 NL 共同激活烟草对烟草花叶病毒的抗性[76]。拟南芥 RPS4 可变剪切产生的其中一条转录本编码一种截短蛋白, 该蛋白能够自激活引发植物细胞坏死[77]。RPS4 通过可变剪切调控多个转录本的表达以抑制自激活, 维持植物正常生长发育。早期鉴定到的抗病基因可变剪切事件大多发生在 TNL 类抗病基因中, 这主要源于 TNL 类基因含有较多内含子结构, 增加了可变剪切发生的可能性。研究发现 CNL 类抗病基因也存在可变剪切现象并且不同转录本在植物免疫中发挥不同的功能。Dennis A. Halterman 等人发现大麦 CNL 基因 Mla13 在上游开放阅读框(uORF) 区域含有两个内含子并通过可变剪切产生五种转录本, 在病原菌侵染过程中,Mla13 内含子剪切转录本转录上调, 激活大麦抗性[78];水稻CNL基因Pi-ta能够发生可变剪切产生 11 种转录本,其中5 个转录本正常表达并编码不同的蛋白, 病原菌侵染时抗病品种中编码 TPX 结构域蛋白的转录本表达水平升高[79];小麦 NLR 基因Lr10 发生可变剪切产生两种转录本, 并且同时含有这两种转录本的硬粒小麦对叶锈病表现抗性[80];水稻抗病基因 RGA5 通过内含子保留形式产生两种转录本,而发生可变剪切的区域包含抗病基因与其无毒基因 Avr1-CO39 识别的关键区段,导致内含子保留转录本编码的抗病蛋白无法识别 Avr1-CO39 [81]。目前,NLR基因的可变剪切调控机制还未被完全解析。Xu等人首次发现植物一类富含丝氨酸-精氨酸蛋白家族(serine/arginine-rich protein, SR)中的MOS12功能缺失会影响拟南芥抗病基因SNC1及RPS4的剪切模式[82]。马铃薯广谱抗病基因 RB的转录本表达量在侵染过程中受其无毒基因AVRblb1调控, 同时发现马铃薯中保守的剪切因子CWC15参与到可变剪切调控过程中进而影响RB介导的晚疫病抗性[83]。无义介导的mRNA 降解(nonsense-mediated mRNA decay,NMD)作为一种对可变剪切产物加工的机制, 通过识别并降解含有提前终止密码子的 mRNA, 避免产生截短的蛋白产物对细胞产生毒性[84]。NMD对抗病基因表达也具有调控作用 研究发现植物中NMD复合物的靶标中近三分之一是抗病基因[85]。例如, NMD突变体中RPS6的内含子剪切转录本会过量积累导致植物生长缺陷[86, 87]。植物小RNA(small RNAs,sRNAs) 参与调控生长发育及胁迫响应等多种生命活动,是基因表达的重要调控因子。sRNA主要分为两类:微小RNA(micro RNA,miRNA)以及小干扰RNA(small interfering RNA,siRNA)。sRNAs主要通过与靶标mRNA序列反向互补抑制基因表达,或通过对mRNA进行切割降解mRNA 或抑制翻译[88]。植物抗病基因的表达水平受到不同 miRNAs及siRNA的调控作用。Yi和Richards首次发现拟南芥中的sRNAs参与调控抗病基因的表达,其研究表明包括21nt siRNA在内的多个siRNA共同作用抑制RPP5基因簇上包括SNC1及 RPP4在内的多个抗病基因的表达水平[89]。其他植物中发现一些miRNA基因家族参与调控抗病基因的表达。例如,蒺藜苜蓿(Medicago truncatula)中三个miRNA 家族靶向多达74个抗病基因,并进一步生成具有相位排列结构特征的植物内源 siRNA(phase secondary small interfering RNA,PhasiRNAs),这类小RNA 通过与AGO蛋白结合对靶标mRNA进行切割降解,最终抑制抗病基因的表达水平[90]。有报道phasiRNAs在豆类及茄科植物中也能够靶向抗病基因,并导致其转录后沉默[91]。因此, 通过编辑植物内源sRNAs调控抗病基因的精细表达,可为平衡抗病与生长发育提供新策略[92]。3 蛋白水平调控
抗病基因在蛋白水平的调控是抗病基因功能调节的重要组成部分,其主要分为蛋白的加工和修饰。加工是指抗病基因翻译后的产物伴随其他蛋白一起组成功能蛋白,这些蛋白质往往称作伴侣蛋白, 而修饰则指通过在氨基酸残基上发生的磷酸化、乙酰化等化学修饰调控抗病蛋白功能。3.1 分子伴侣和辅助伴侣分子调控
与动物中的抗病蛋白相似,植物抗病蛋白也需要其他蛋白来促进其折叠。一类热休克蛋白能够调控蛋白质成熟以及降解错误折叠的多肽,因而影响多个信号转导蛋白的活性,这种蛋白也被成为伴侣蛋白[93]。植物伴侣蛋白HSP90参与调控多个抗病蛋白的稳定性,HSP90的突变或沉默降低了抗病蛋白Rx、RPM1以及RPS5的蛋白表达水平[94-96]。同时,遗传分析表明HSP90介导的蛋白稳定性直接参与调控抗病蛋白介导的免疫反应[97]。此外,研究发现植物保守的伴侣蛋白RAR1和SGT1能够与HSP90发生互作,形成复合体参与调节多个抗病蛋白的稳定性。同时突变 HSP90、SGT1以及RAR1时,包括 N,RPS2在内的多个抗病蛋白表达下降,导致免疫响应降低[98]。单独突变RAR1时,拟南芥RPM1和RPS5、大麦Mla1和Mla6以及马铃薯Rx等多个抗病蛋白的表达均受到影响[96, 99-101]。同样,SGT1的缺失也造成多个抗病蛋白的积累下降, 如Rx以及N[102-104]。
3.2 泛素化调控机制
蛋白质泛素化是一种常见的蛋白质翻译后修饰(Post-translational modification),主要通过泛素蛋白酶体系统(ubiquitin proteasome system, UPS)调控蛋白的降解进而影响其功能。植物抗病蛋白受到复杂的泛素化调控, 以确保激活后的抗病蛋白在发挥抗病功能后被正确降解,以避免过高的抗病反应抑制生长发育。目前,已有的研究表明E3泛素连接酶是调控植物抗病蛋白泛素化的主要蛋白, E3连接复合酶体SCF(SKP1/Cullin1/F-box)中的F-box蛋白CPR1调控拟南芥抗病蛋白SNC1、RPPS2和SUMM2的蛋白降解,当其缺失时会导致这些蛋白水平持续积累[33, 105, 106]。此外,拟南芥中抗病蛋白SIKIC2的蛋白质水平受到两个E3连接酶 MUSE1以及MUSE2的负调控[107]。番茄E3泛素连接酶SBP1与番茄抗病蛋白Sw-5b 互作并介导其降解[108]。烟草E3泛素连接酶UBR7与抗病N蛋白互作,UBR7表达下调导致N蛋白表达升高进而增强烟草对 TMV的抗性水平[109]。尽管目前对抗病蛋白的泛素化修饰的研究取得了很大进展,但鉴于抗病蛋白的分子量较大,蛋白结构复杂,更多翻译后修饰研究在抗病蛋白上的作用机制还没有明确报道,因此挖掘抗病蛋白发生的修饰并解析其功能机制对理解抗病蛋白的作用机制有重要意义,对更好的应用抗病蛋白提供理论基础。
蛋白磷酸化修饰是调控细胞信号转导的主要机制,其通过蛋白激酶的催化作用将磷酸基团转移到蛋白质的氨基酸残基上,如丝氨酸、苏氨酸以及酪氨酸[110]。蛋白磷酸化在PTI免疫过程中发挥重要作用。当PRR受体识别到PAMP信号分子后,与共受体激酶BAK1互作并相互磷酸化,以迅速激活下游免疫反应[111]。研究发现磷酸化对抗病蛋白的激活具有重要意义, 例如,拟南芥抗病基因对RRS1和RPS4分别可以识别青枯雷尔氏菌分泌的效应因子PopP2以及丁香假单胞效应分子AvrRPS4,激活植物对不同病原菌的抗性[112-114]。RRS1在拟南芥中存在等位变异,其中RRS1-S 仅识别AvrRPS4,而RRS1-R可同时识别AvrRPS4和PopP2。序列分析发现,相较于 RRS1-S,RRS1-R末端第1214位苏氨酸可发生磷酸化,且该磷酸化可抑制其自身活性。PopP2编码一个乙酰转移酶,可将RRS1- R末端磷酸化位点乙酰化,从而解除对 RRS1-R的抑制,激活下游免疫响应[115]。该研究解析了不同蛋白修饰对同一抗病蛋白活性的调控机制,同时也展示了一个植物抗病蛋白在进化上利用自身蛋白修饰位点诱捕病菌效应蛋白的新策略。
抗病蛋白的亚细胞定位与其功能息息相关。目前,已报道的抗病蛋白主要在细胞膜、细胞质及细胞核中发挥功能。研究发现,部分抗病蛋白具有质核定位的变化, 并且该变化对于抗病蛋白的激活具有重要作用[116]。例如, 拟南芥SNC1蛋白在细胞核中积累会造成免疫自激活,而将核输出信号与 SNC1 融合后可抑制SNC1 的自激活, 表明核定位对于SNC1介导的免疫激活至关重要[117-119]。进一步研究发现, SNC1的入核受到输入蛋白 MOS6 的调控, 而 SNC1 的出核受转运蛋白 KA120 调控[120, 121]。部分抗病蛋白在不同亚细胞结构中均发挥功能, 但其作用机制存在差异。番茄抗病蛋白 Sw-5b 分布于细胞质和细胞核中,研究发现定位在胞质中有利于 Sw-5b 诱导强烈的细胞死亡但无法阻止病毒在细胞间和长距离的移动;而定位在细胞核中的 Sw-5b 虽触发的免疫较弱,但能有效阻止病毒在细胞间和全系统中的扩散。Sw-5b 定位的变化受到其 N 端 SD 结构域与转运蛋白互作的调控, 表明植物抗病蛋白能够通过核质穿梭调控植物抗性水平[122]。此外,植物抗病蛋白激活后形成抗病小体发挥功能,下游 helper NLR 蛋白 NRG1 和ADR1 可在细胞膜上形成钙离子通道,激活下游免疫响应,同时,也有研究发现NRG1 和 ADR1 在核里与信号蛋白 EDS1 形成复合体,参与抗病信号的传递[123- 126]。虽然抗病蛋白亚细胞定位与功能的联系已得到广泛验证,但是其分子机制尚不清晰。除核孔转运蛋白外,核孔复合体蛋白、 蛋白修饰以及蛋白序列的变化均可能参与抗病蛋白亚细胞定位的变化[127-129]。未来对抗病蛋白调控机制的深入研究有望解析其亚细胞定位变化的调控机理,并为抗病品种的培育提供新思路。