南京师范大学开发转录因子代谢重编程策略高效生产DHA
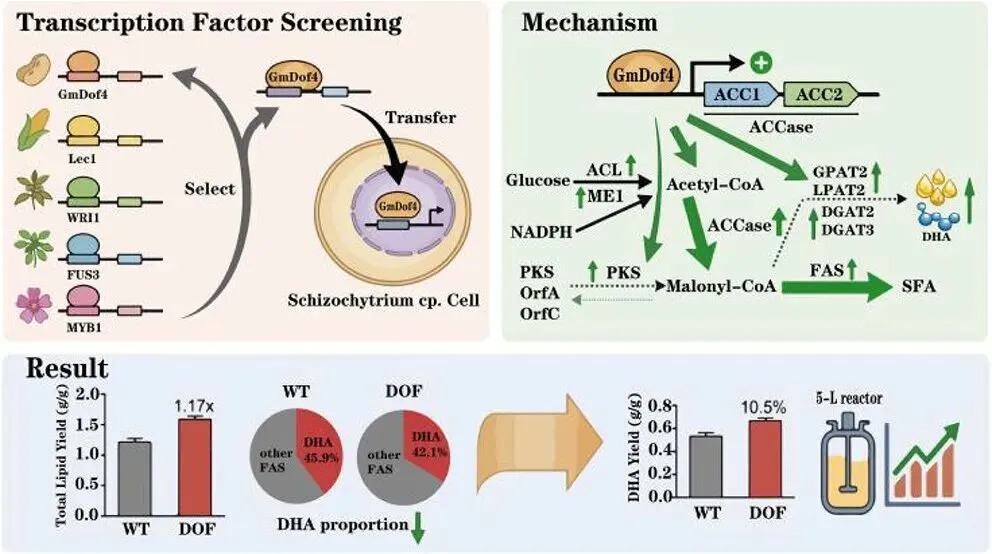
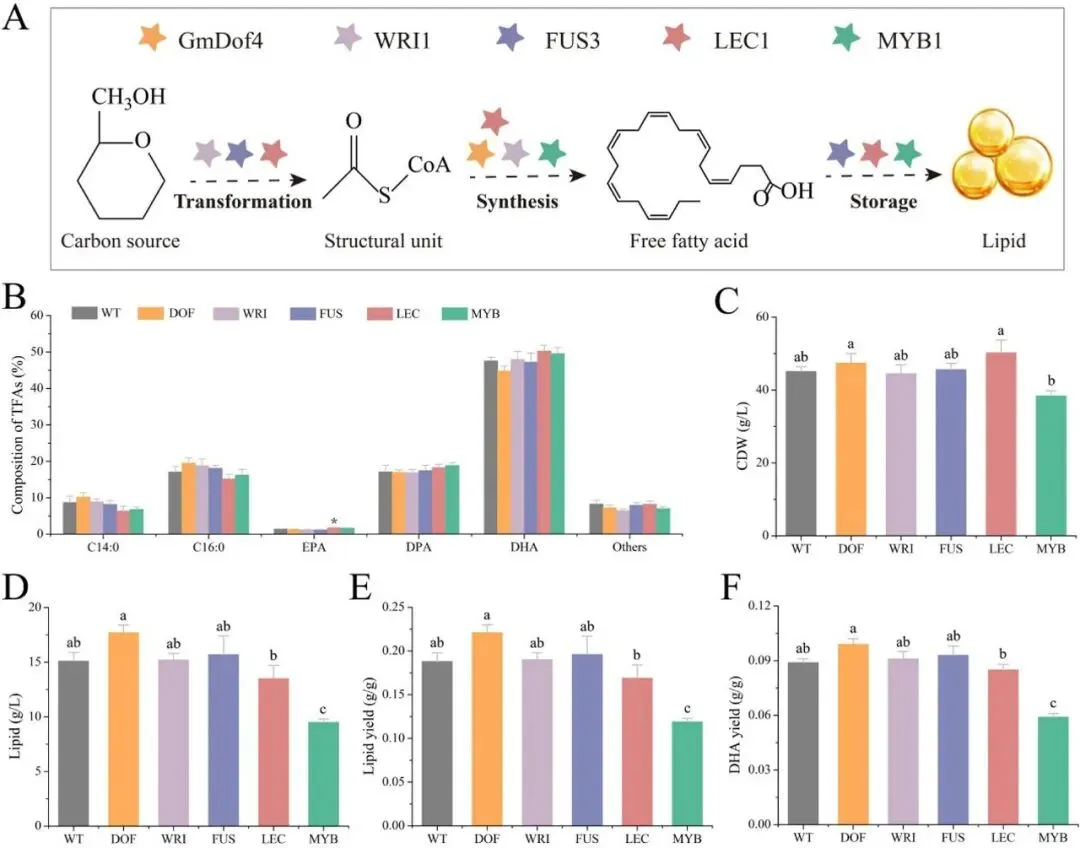
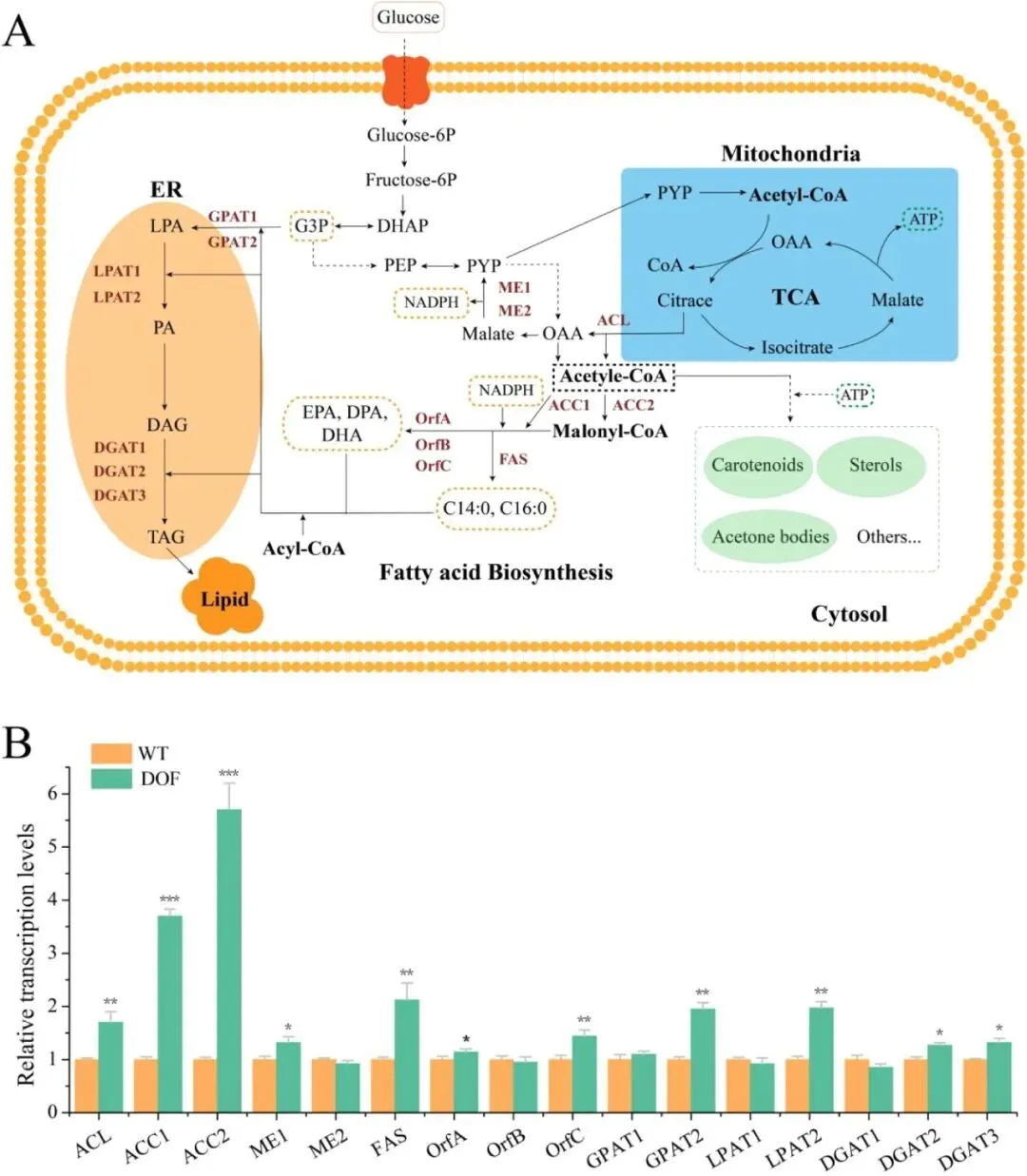
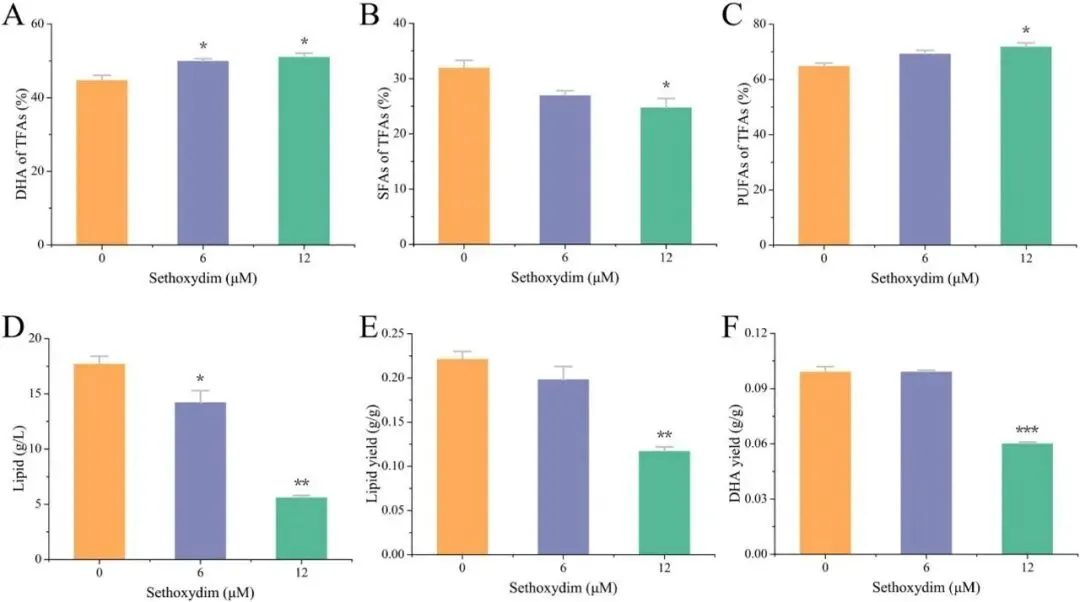
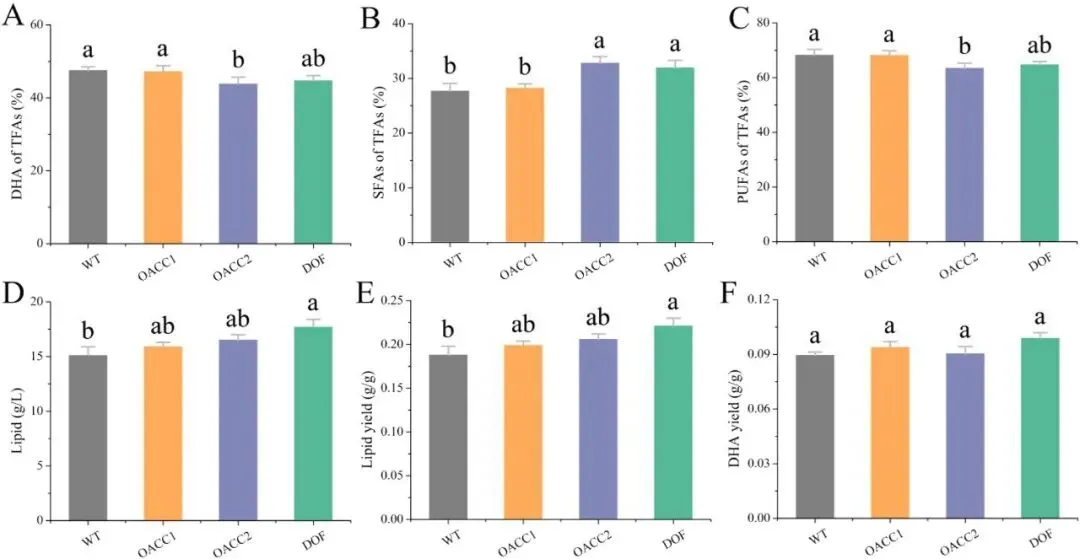
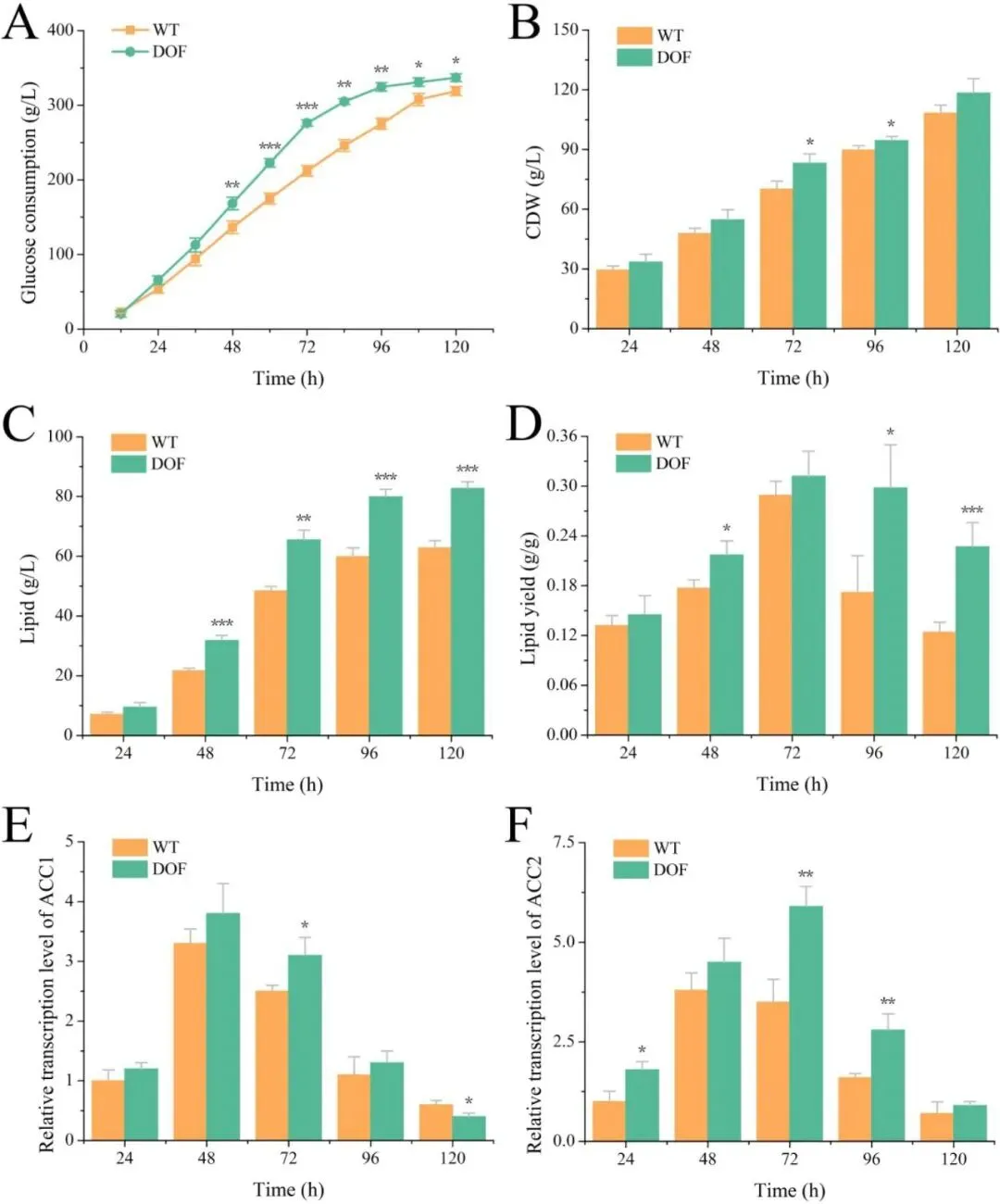
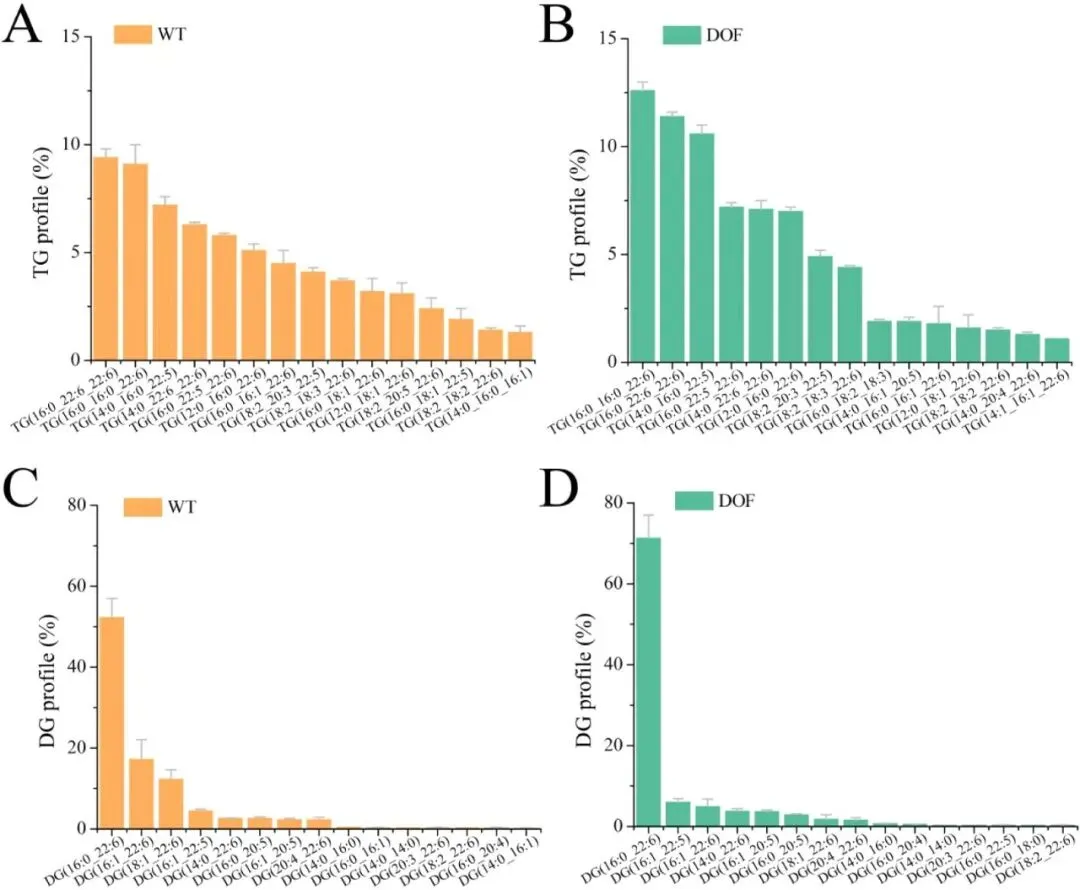
二十二碳六烯酸(DHA)是维持人体大脑、视网膜及心血管健康的必需多不饱和脂肪酸,利用海洋产油微生物裂殖壶菌(Schizochytrium sp.)发酵生产 DHA 油脂已成为替代传统鱼油的主流可持续方案。然而,脂质产率作为碳源转化效率的核心指标,始终是制约该过程经济性的关键瓶颈。传统代谢工程多采用自下而上策略,直接改造脂质合成途径中个别酶的活性,但单点强化易导致代谢通量失衡、中间产物积累或细胞生长胁迫,难以实现全局碳流的最优重定向。2026年7月9日,来自南京师范大学的马旺教授团队在《Journal of Agricultural and Food Chemistry》杂志上发表了题为“Systematic Metabolic Rewiring of Schizochytrium sp. via GmDof4 Overexpression for Enhanced DHA-Rich Lipid Production”的研究论文。该研究采用自上而下的系统工程策略,从五种异源转录因子中筛选出大豆来源的 GmDof4 作为最强效调控因子,通过其在裂殖壶菌中的过表达实现了脂质合成网络的全局协同调控。在 5 L 生物反应器补料分批发酵中,工程菌株总脂质产量达到 82.7 g/L,较野生型提高 31.7%,葡萄糖转化效率提升 22.4%;尽管 DHA 相对比例从 45.9% 降至 42.1%,但其绝对产量仍达到 34.8 g/L,增幅达 20.8%,DHA 产率提高 14.4%。机制研究表明,GmDof4 主要通过显著上调乙酰辅酶 A 羧化酶(ACCase)两个亚型 ACC1 和 ACC2 的表达(分别提升 2.7 倍和 4.7 倍),同时协调增强前体供应、脂肪酸合成及三酰甘油组装等多个代谢阶段,实现了代谢通量的系统性重分配。图1. GmDof4 介导的裂殖壶菌脂质合成转录调控机制概念图研究人员首先筛选了五种异源转录因子(GmDof4、WRI1、FUS3、LEC1、MYB1)对裂殖壶菌发酵性能和脂肪酸组成的影响,以鉴定最具脂质增强潜力的调控因子。结果显示,不同转录因子的调控效应存在显著差异:LEC1 菌株使 EPA 比例上升且 DHA 比例提高 5.8%,生物量增加 11.2%,但脂质产量仅为野生型的 89.5%;MYB1 菌株使 EPA 和 DHA 比例分别提升 18.6% 和 4.3%,但脂质产率和 DHA 产率分别下降 36.9% 和 34.3%;WRI1 和 FUS3 菌株与野生型相比无显著差异。相比之下,GmDof4 过表达菌株(DOF)表现出最优的综合生产性能,其脂质产率达到 0.221 g/g,为野生型的 1.17 倍;尽管 DHA 相对比例下降 6.1%,但 DHA 绝对产率仍达 0.099 g/g,较野生型提高 10.5%。这一结果表明,总脂质合成能力的增强足以抵消 DHA 比例的下降,最终实现更高的 DHA 绝对产量,因此 GmDof4 被确定为本研究筛选出的最具应用潜力的候选转录因子。为揭示 GmDof4 提升裂殖壶菌脂质产量的分子机制,研究人员系统比较了野生型与 DOF 菌株中脂质合成相关关键基因的转录水平。结果表明,GmDof4 并非非选择性地全局激活脂质代谢网络,而是呈现高度靶向的层级调控模式,其核心作用是显著上调脂肪酸从头合成的限速步骤。其中,编码 ACCase 两个亚型的 ACC1 和 ACC2 基因表达分别提升约 2.7 倍和 4.7 倍,上调幅度最为显著;提供乙酰辅酶 A 前体的 ATP - 柠檬酸裂解酶(ACL)基因表达上调 70%;负责 NADPH 供应的苹果酸酶 ME1 转录水平提升 32%;脂肪酸合酶 FAS 的表达也被同步激活,达到野生型的 2.1 倍。在下游三酰甘油组装途径中,GPAT2 和 LPAT2 分别上调 95% 和 97%,而 DGAT2 和 DGAT3 仅分别上调 27% 和 32%,GPAT1、LPAT1 和 DGAT1 则无显著变化。对于 DHA 合成关键的 PKS 系统基因 OrfA 和 OrfC,仅分别上调 14% 和 44%。这种上游强化显著强于下游、FAS 途径激活远强于 PKS 途径的调控模式,直接解释了总脂质大幅增加而 DHA 相对比例下降的表型。图3. 转录因子 GmDof4 对裂殖壶菌发酵指标的影响分析为验证 ACCase 在 GmDof4 介导的脂质增产效应中的核心作用,研究人员采用 ACCase 特异性抑制剂烯禾啶(sethoxydim)对 DOF 菌株进行处理。结果显示,随着抑制剂浓度从 0 μM 增加至 12 μM,DOF 菌株的脂肪酸组成发生系统性变化:DHA 占总脂肪酸的比例从 44.7% 升至 51.0%,多不饱和脂肪酸总比例从 64.7% 升至 71.7%,而饱和脂肪酸比例则从 31.9% 降至 24.7%。与此同时,菌株的生长和脂质合成均受到剂量依赖性抑制:无抑制剂时 DOF 菌株的脂质产率和 DHA 产率分别为 0.221 g/g 和 0.099 g/g,而在 12 μM 烯禾啶处理下,脂质产率降至 0.117 g/g,DHA 产率降至 0.060 g/g,DOF 菌株相对于野生型的生产优势基本消失。该结果直接证明,ACCase 活性是 GmDof4 发挥代谢重编程效应的必要前提,抑制该酶的功能可大幅削弱转录因子介导的脂质增产效果。为进一步判断 ACCase 上调是否为 GmDof4 功能的唯一机制,研究人员分别构建了 ACC1 和 ACC2 单基因过表达菌株(OACC1、OACC2),并与 DOF 菌株进行系统比较。结果显示,OACC1 和 OACC2 菌株的脂质产率和 DHA 产率均较野生型有所提升,证实 ACCase 活性增强本身即是提高脂质积累的有效策略,其中 OACC2 菌株的脂质产率略高于 OACC1,但其 DHA 比例降至 43.8%,DHA 产率表现欠佳。然而,与单独过表达 ACC1 或 ACC2 的菌株相比,DOF 菌株在总脂质产量、脂质产率和 DHA 产率等关键指标上均表现更优。尽管 DOF 菌株的 DHA 比例略低于野生型和 OACC1 菌株,但其更高的总脂质产量最终带来了最高的 DHA 绝对产量。更重要的是,DOF 菌株呈现出独特的脂肪酸谱,其饱和脂肪酸与多不饱和脂肪酸的比例异于野生型及其他工程菌株,表明 GmDof4 的调控范围超出 ACCase 本身,涉及更广泛的脂质代谢网络。综合抑制与增强实验的结果表明,ACCase 是 GmDof4 不可或缺的功能靶点,但其完整的调控效应无法通过单一酶的过表达完全复制。为评估 GmDof4 转录因子工程的规模化应用潜力,研究人员在 5L 生物反应器中对野生型和 DOF 菌株进行了补料分批发酵。结果表明,DOF 菌株在整个发酵过程中均表现出更优的细胞生长和底物利用效率:发酵前期(24-72 h)葡萄糖消耗速率显著高于野生型;发酵中期 48 h 和 72 h 时细胞干重分别达到 54.7 g/L 和 83.2 g/L,显著高于同期野生型的 47.9 g/L 和 70.2 g/L。脂质产量方面,DOF 菌株从 48 h 起持续领先,发酵 120 h 时脂质终产量达 82.7 g/L,较野生型的 62.8 g/L 提高 31.7%;脂质产率达 0.245 g/g,较野生型的 0.196 g/g 提升 22.4%,表明其底物转化效率的实质性提高而非单纯消耗更多底物。转录动力学分析显示,在脂质快速积累阶段(24-72 h),DOF 菌株中 ACC1 和 ACC2 的转录水平始终高于野生型,分别在 48 h 和 72 h 达到峰值,与脂质积累速率的变化趋势高度吻合,从时间维度进一步支持了 GmDof4 通过持续上调 ACCase 表达驱动脂质过量合成的核心机制。图6. 野生型与 DOF 工程菌株补料分批发酵性能指标为在分子层面系统评估 GmDof4 对裂殖壶菌脂质组成的影响,研究人员对野生型和 DOF 菌株发酵产物进行了非靶向脂质组学分析,重点考察三酰甘油(TG)和二酰甘油(DG)的脂肪酸分布。结果显示,两菌株的优势 TG 物种发生显著重构:野生型中含量最高的 TG (16:0-22:6-22:6) 占比为 9.4%,含两个 DHA 分子的 TG 物种占主导;而 DOF 菌株中 TG (16:0-16:0-22:6) 升至首位,相对含量大幅增至 12.6%,TG (16:0-22:6-22:6) 则退居第二位且仅为 11.4%。DG 作为 TG 合成的直接前体,其组成变化更为显著:两菌株中含量最高的均为 DG (16:0-22:6),但其在野生型中占比为 52.3%,在 DOF 菌株中则大幅升至 71.3%,同时其他含 DHA 的 DG 物种比例显著下降。这些结果表明,GmDof4 引发了裂殖壶菌脂质代谢的系统性重编程,使 TG 分布从富含 DHA 的模式向富含 C16:0-DHA 的构型转变,反映出上游脂肪酸合成通量扩增与 PKS 途径相对滞后之间的通量不对称分配。总之,本研究通过在裂殖壶菌中异源表达大豆转录因子 GmDof4,成功实现了脂质生物合成网络的自上而下系统性重编程,验证了转录因子工程作为构建高效微生物 DHA 生产平台的强大策略价值。该研究不仅阐明了 GmDof4 以 ACCase 为核心、多靶点协同调控的作用机制,还在 5L 生物反应器规模上实现了总脂质产量 31.7% 和 DHA 绝对产量 20.8% 的显著提升,为降低 DHA 发酵生产成本、推动其工业化应用提供了新的技术路径。这一成果突破了传统单点代谢工程的局限,为产油微生物的全局代谢优化提供了新思路,未来可结合 PKS 途径的协同调控进一步提升 DHA 比例,构建性能更优的下一代 DHA 生产菌株。