📌 摘要
本研究系统探究了聚苯乙烯纳米塑料(PS NPs,粒径为5.4±1.0 nm)对水稻(Oryza sativa L.)光合作用的影响。通过为期14天的水培实验,比较了根部暴露(3.6-35.7 μg PS/株/天)与叶面暴露的效应,并结合为期30天的土壤栽培叶面暴露实验(从抽穗期至成熟期,5.3-73.4 μg PS/株/天)。研究发现,PS NPs的植物毒性具有显著的途径依赖性:仅高剂量的根部暴露会抑制光合作用,而叶面暴露则会引发持续性的光合抑制。更重要的是,在土壤栽培的叶面暴露实验中,即使PS NPs未转运至稻谷中,其对光合作用的抑制依然发生。这种叶面特异性抑制与PS在叶片中的积累量直接相关,其机制在于PS与叶绿体的共定位(Pearson's r = 0.592),这在共聚焦显微镜下得到证实。这种共定位损害了光合作用的光反应和暗反应,具体表现为光合色素显著减少(54.4-61.0%)、希尔反应活性降低(49.9-70.2%)、ATP产量下降(22.1-24.3%)、净光合速率锐减(84.5-89.0%)、Rubisco酶活性降低(84.6-87.5%)、气孔导度(47.9-69.5%)和蒸腾速率(47.4-48.9%)下降。转录组学分析揭示了其遗传基础,表明核心光合作用途径、光合天线蛋白、碳固定以及类胡萝卜素/卟啉代谢的关键基因均显著下调。这些发现为PS NPs诱导的光合抑制提供了机制性见解,并强调大气纳米塑料是对全球粮食安全的新兴威胁。
🧭 引言
纳米塑料(NPs)在全球范围内无处不在,从陆地到大气,存在于各种环境中。植物可以通过根系和叶面两种途径吸收NPs,从而引发植物毒性,包括生长抑制、形态异常、生理功能障碍、氧化应激和基因毒性。值得注意的是,NPs会降低光合色素(即叶绿素a/b)的含量。我们前期结合了157篇文献的荟萃分析和机器学习方法的研究表明,微/纳米塑料暴露会显著降低植物叶绿素含量(5.63-17.42%),这相当于导致全球主要农作物每年损失1.0973-3.6087亿吨的产量。尽管这些研究确凿地证明了NPs诱导的光合抑制,但几个关键的知识空白仍然存在。
首先,目前缺乏在环境相关浓度下,对根部暴露与叶面暴露导致的光合抑制进行系统且等效的比较,这严重阻碍了我们阐明途径特异性效应的能力。当前研究存在显著的不平衡:截至2025年12月,94%的发表文献不成比例地优先研究根部暴露,而忽视了叶面暴露(附录列表S1)。然而,越来越多的证据强调了叶面暴露的生态相关性。实地证据证实,陆地植物叶片会吸收并积累空气中的微/纳米塑料,其中聚苯乙烯(PS)和聚对苯二甲酸乙二醇酯(PET)等常见聚合物的浓度在露天蔬菜中比温室蔬菜高出一到两个数量级,并且在外叶中的含量高于新生叶。尽管叶面暴露意义重大,但在研究中仍被严重忽视,截至2025年底仅占发表文献的6%(附录列表S1)。鉴于全球大气样本中微/纳米塑料的检测频率越来越高(浓度范围为10⁻²至10³个/m³),这种研究差距尤其令人担忧。据估计,美国受保护地区的大气微/纳米塑料年沉降量超过1000公吨,而中国长江流域湿地则达到17.46公吨。这些沉降的颗粒物以每平方厘米叶片0.07至0.19个颗粒的丰度积累在植物叶片表面,从而为包括水稻在内的主要湿地作物建立了一条暴露途径。因此,理解这些途径特异性效应对于阐明暴露途径在光合毒性中的作用,以及为制定农业和自然生态系统中的针对性法规和有效缓解策略至关重要。
其次,一个关键的挑战是将NPs的物理存在和细胞内定位与其毒性机制联系起来,特别是与光合作用器联系起来。研究表明,PS NPs的叶面吸收取决于粒径和表面电荷,更小、带正电的NPs在叶片组织中表现出更高的积累。这种生物积累不仅通过降低叶绿素含量(8.5-34.8%)和光合能力(9.2-27.9%)导致生物量减少(14.3-23.2%),还会诱导氧化应激并破坏氨酰-tRNA生物合成。这些不利影响可在营养级间传播:在生菜-蜗牛食物链中,积累的PS NPs降低了蜗牛的适口性并破坏了其消化系统。理解PS NPs抑制光合作用的分子机制是一个关键步骤。特别是,NP亚细胞定位与其导致的光合功能损伤之间的直接联系仍然难以捉摸。
本研究旨在通过以下三个方面探究PS NPs对水稻(Oryza sativa L.)光合作用的影响:(1)比较根部暴露与叶面暴露后的光合响应;(2)探究PS积累与光合响应之间的相关性;(3)研究光合作用抑制的分子机制。PS NPs同时标记了荧光素和钯,这使得能够同时进行NPs的定量和高分辨率追踪。这些结果为评估NPs对农业生产的潜在风险提供了宝贵的见解。
🧪 材料与方法
PS NPs的合成与表征:
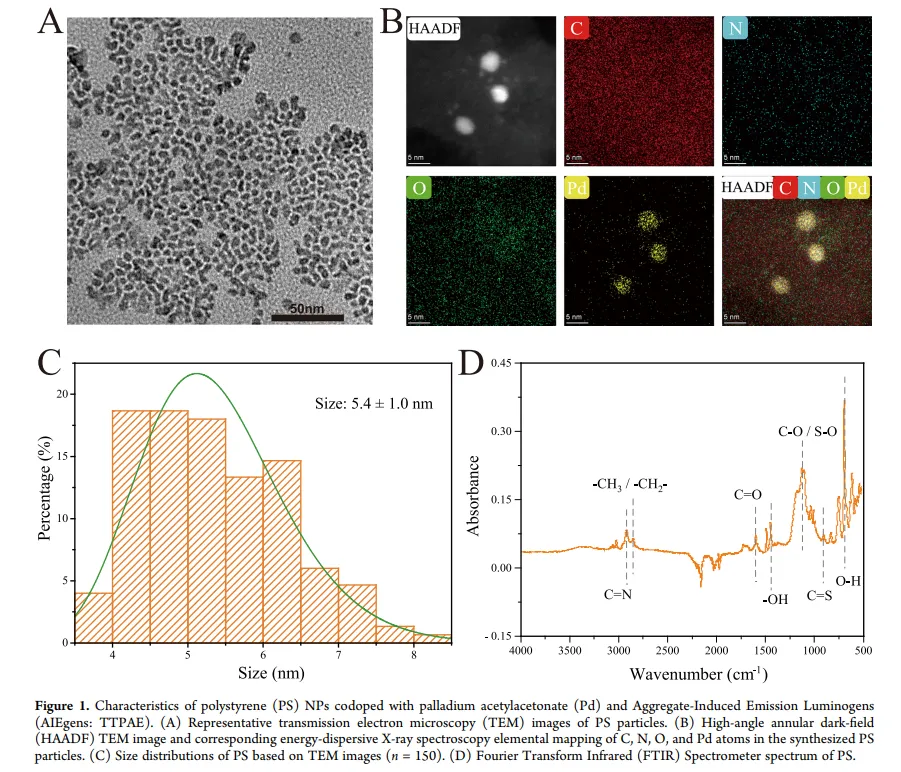
通过乳液聚合法合成了共掺杂乙酰丙酮钯(Pd)和聚集诱导发光染料(AIEgens)的PS NPs,粒径为5.4 ± 1.0 nm,Zeta电位为 -49.1 ± 0.80 mV。
使用透射电子显微镜(TEM)、高角环形暗场扫描TEM(HAADF-STEM)结合能谱(EDS)进行形貌和元素分析。
通过傅里叶变换红外光谱(FTIR)确认其表面化学性质为PS。
示踪剂浸出实验表明,荧光素和Pd的浸出率极低(<0.3%和<0.09%),证实了其作为PS保守示踪剂的可靠性。
PS与Pd浓度呈线性相关,用于定量分析。
水稻暴露实验:
水培实验(14天):比较根部暴露与叶面暴露。设置对照组、低剂量组(3.6 μg PS/株/天)和高剂量组(35.7 μg PS/株/天)。每组3个盆,每盆4株。
土壤栽培实验(30天):模拟田间条件,仅进行叶面暴露(从抽穗期至成熟期),剂量为5.3和73.4 μg PS/株/天。每组5个盆,每盆3株。
暴露方式:叶面暴露时,使用Silwet L-77辅助PS NPs均匀沉积于叶片表面,每日喷雾。根部暴露则将水稻根系置于含PS NPs的营养液中。
收获后,植物组织经过严格清洗以去除表面吸附的PS NPs。
PS NPs在植物体内的定位与定量:
光合作用参数测定:
转录组学分析:
📊 结果
PS NPs在水稻体内的积累与转运具有途径依赖性
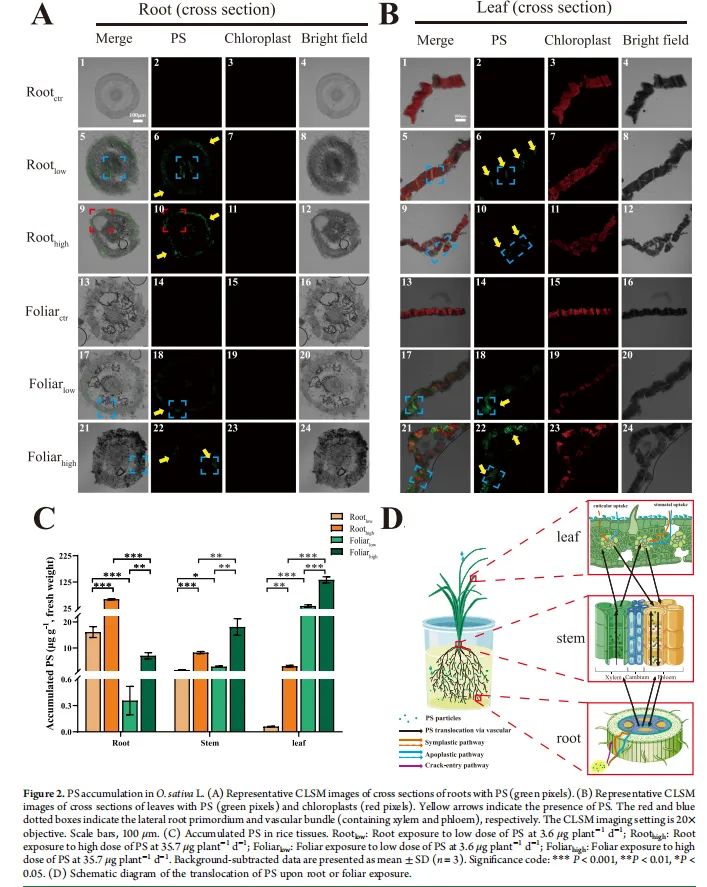
可视化证据:CLSM图像显示,根部暴露后,PS NPs(绿色荧光)定位于根尖、侧根原基和维管束(木质部),并可向上转运至茎和叶。叶面暴露后,PS NPs通过气孔进入叶片,定位于维管束(韧皮部),并可向下转运至茎和根。
定量证据:PS积累量随暴露剂量增加而增加。在同等暴露剂量下,叶面暴露导致茎和叶中的PS浓度分别是根部暴露的1.8-2.2倍和45.0-564.4倍;而根部暴露导致根中的PS浓度是叶面暴露的8.5-45.1倍。
转运机制:根部到叶片的转运主要通过木质部(向上),而叶片到根部的转运主要通过韧皮部(向下)。在为期30天的土壤栽培叶面暴露实验中,尽管叶片中积累了大量的PS(8.86-38.21 μg/g),但未检测到其向稻谷中的转运。
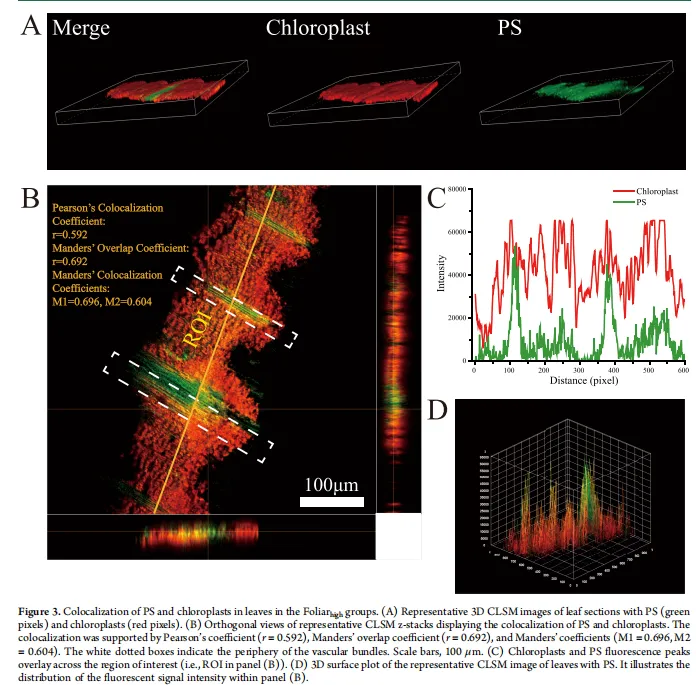
叶面暴露导致PS NPs与叶绿体共定位
3.PS NPs抑制光合作用具有途径和剂量依赖性
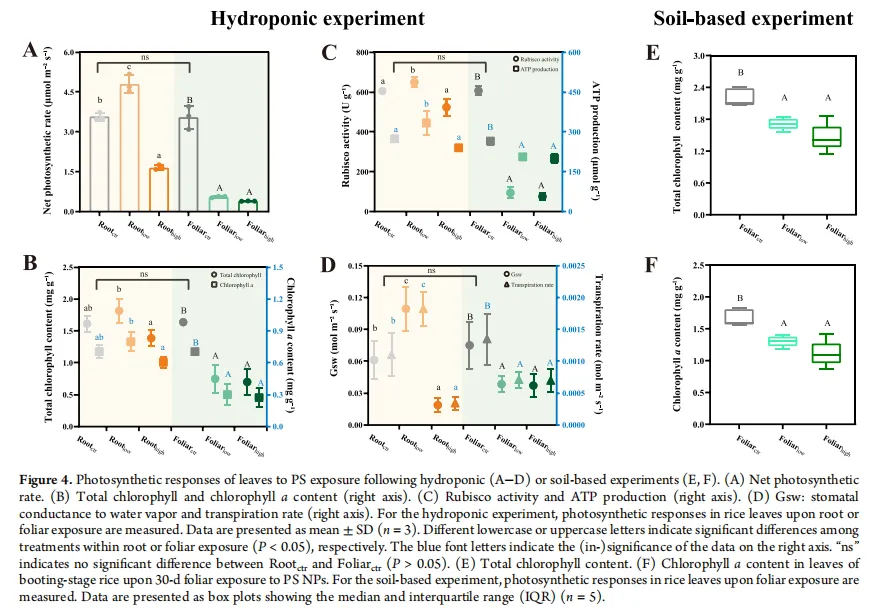
叶面暴露:在所有剂量下均显著抑制光合作用。与对照组相比,净光合速率下降84.5-89.0%,总叶绿素含量下降54.4-56.9%,Rubisco活性下降84.6-87.5%,ATP产量下降22.1-24.3%,气孔导度和蒸腾速率也显著降低。希尔反应活性下降49.9-70.2%,表明光反应电子传递链受损。这些效应在土壤栽培实验中得到验证。
根部暴露:表现出剂量依赖性的双相效应。低剂量下,净光合速率、气孔导度和蒸腾速率反而增加(分别增加34.4%、66.0-78.9%),呈现“毒物兴奋效应”。高剂量下,这些参数才显著降低(净光合速率下降53.6%)。
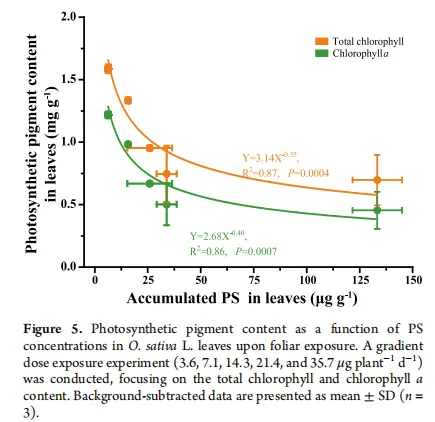
相关性分析:叶面暴露下,叶片中PS的积累量与总叶绿素和叶绿素a含量呈显著负相关,建立了剂量-效应关系。
4.转录组学揭示了光合作用抑制的分子机制
光合作用通路:涉及光系统II(Psb家族,如 psbA, psbB)、光系统I(Psa家族,如 psaA, psaB)、细胞色素b6/f复合体和光合电子传递链的多个关键基因均显著下调。
光合天线蛋白:编码捕光复合物LHC I和LHC II的多个基因(如 Lhca, Lhcb 家族)表达显著下调。
碳固定:编码Rubisco活化酶(RCA)、Rubisco小亚基(RBCS)等关键酶的基因表达下调。
色素代谢:编码叶绿素合成(如 hemA, chlD, chlI)和类胡萝卜素合成(如 PSY, PDS)途径的关键酶基因表达也受到抑制。
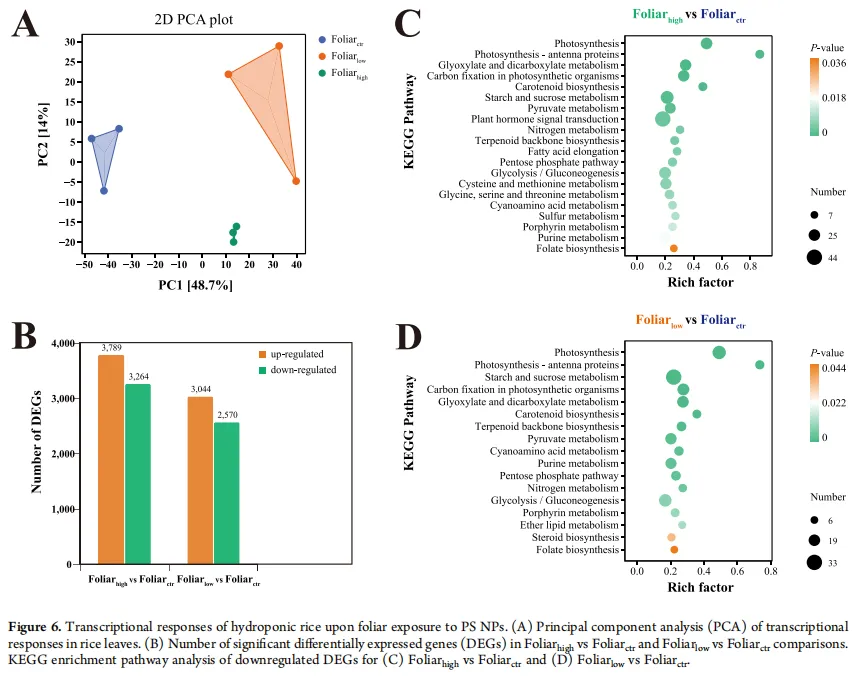
差异表达基因(DEGs):叶面高剂量组(Foliarhigh vs 对照)有1919个DEGs(1244个下调,675个上调),低剂量组(Foliarlow vs 对照)有225个DEGs(120个下调,105个上调)。
KEGG富集分析:下调的DEGs显著富集在光合作用、光合天线蛋白、碳固定、卟啉与叶绿素代谢和类胡萝卜素生物合成等关键通路上。
氧化应激响应:抗氧化酶基因(如超氧化物歧化酶 SOD、过氧化氢酶 CAT 家族)显著上调,表明PS NPs诱导了氧化应激。
综上所述,本研究首次系统阐明了PS NPs通过不同暴露途径对水稻光合作用的差异性抑制效应。其核心机制在于,叶面暴露的PS NPs能够直接进入叶片并与叶绿体共定位,从而在物理和基因层面双重干扰光反应和暗反应的正常运行,最终导致光合效率的全面下降。这一发现将大气纳米塑料污染确认为一个不容忽视的农业生态风险因子。
如有侵权,请联系删除。

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
