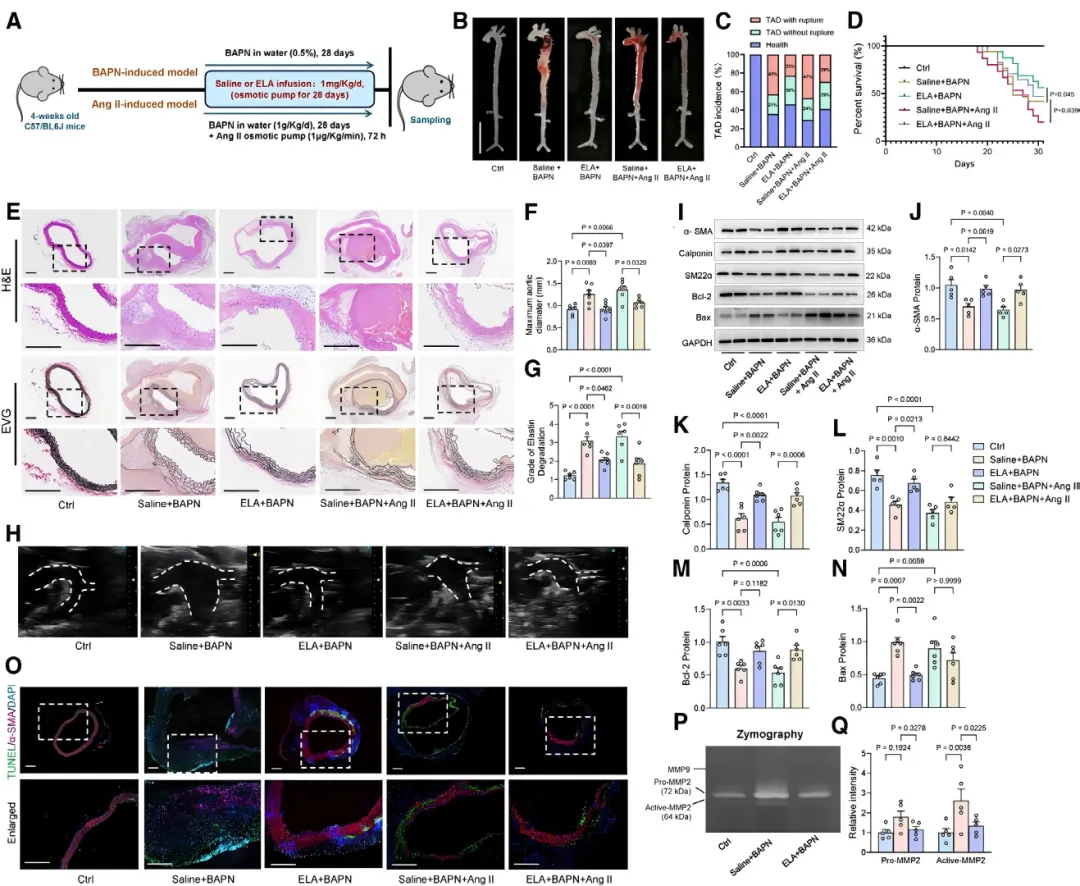
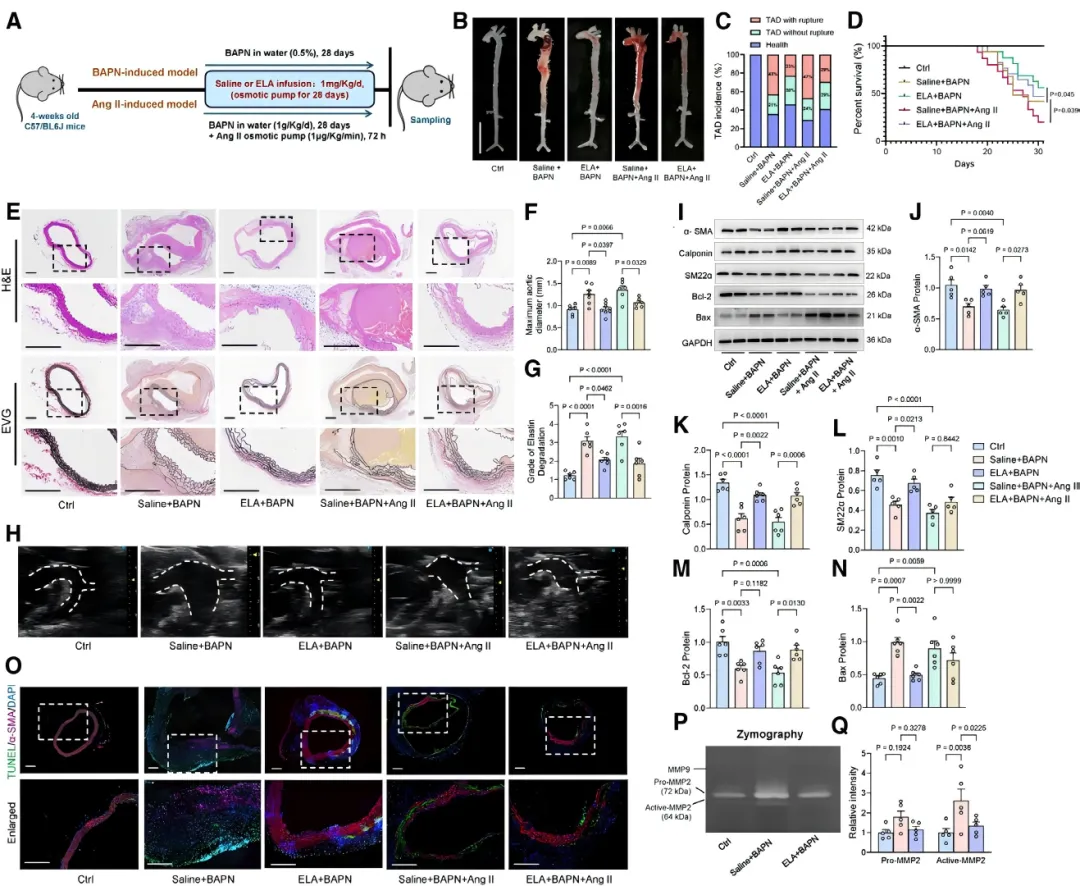
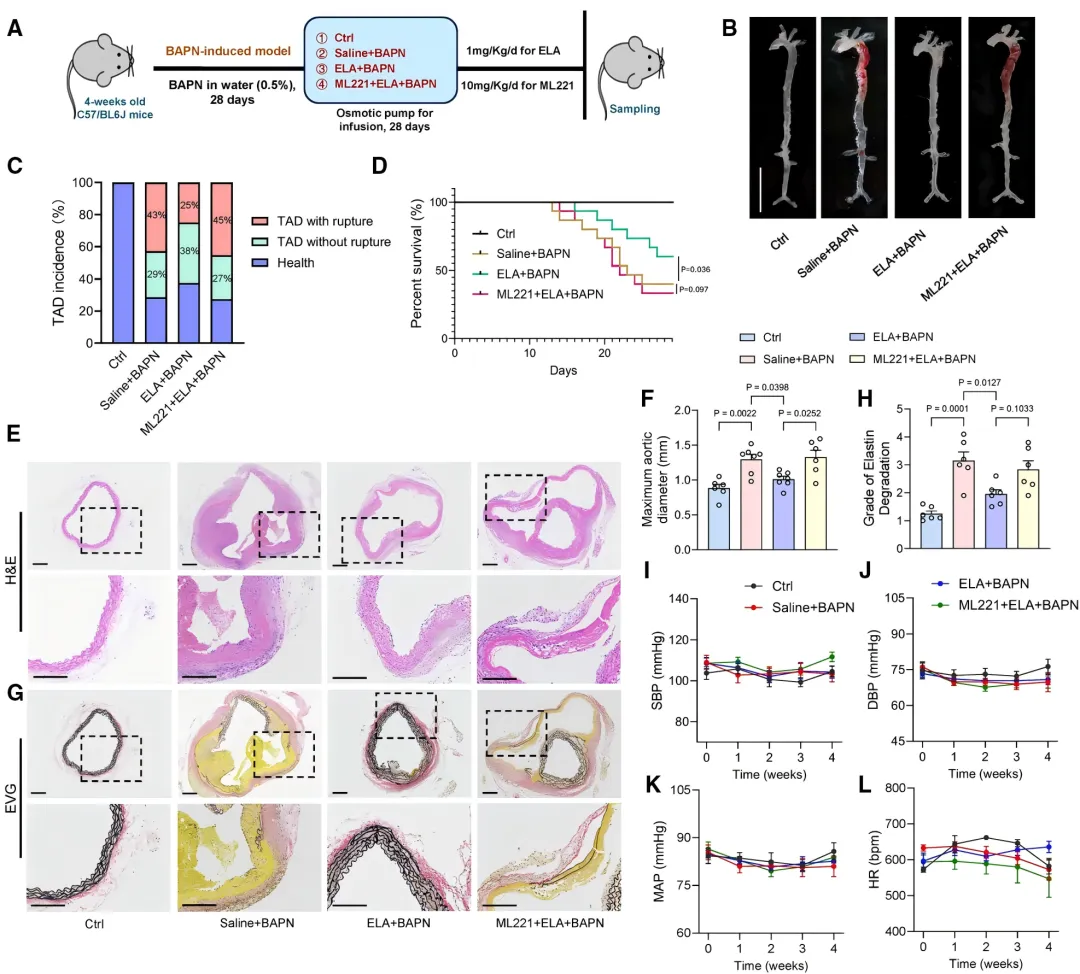
本研究首次揭示了ELA在胸主动脉夹层(TAD)中的保护作用。在两种经典的TAD小鼠模型中,持续输注ELA肽均可显著降低小鼠的死亡率【图1D】。
图1.输注ELA(ELABELA)
可减轻小鼠胸主动脉夹层(TAD)的形成,
并维持血管平滑肌细胞(VSMC)稳态
通过主动脉大体标本【图1B】、超声及组织学染色【图1E, 1H】观察发现,ELA治疗有效抑制了主动脉扩张,并减轻了血管中膜弹性纤维降解【图1F, 1G】,证实ELA能有效延缓TAD病理进程。
TAD的关键病理特征是血管平滑肌细胞(VSMC)稳态失衡。研究发现,ELA输注显著提升了TAD小鼠主动脉中收缩型VSMC标志蛋白的表达【图1I-1L】,同时减少了细胞凋亡【图1M-1O】。
图1.输注ELA(ELABELA)
可减轻小鼠胸主动脉夹层(TAD)的形成,
并维持血管平滑肌细胞(VSMC)稳态
此外,ELA还抑制了基质金属蛋白酶活性【图1P, 1Q】,防止细胞外基质降解,从而保护主动脉壁结构完整。
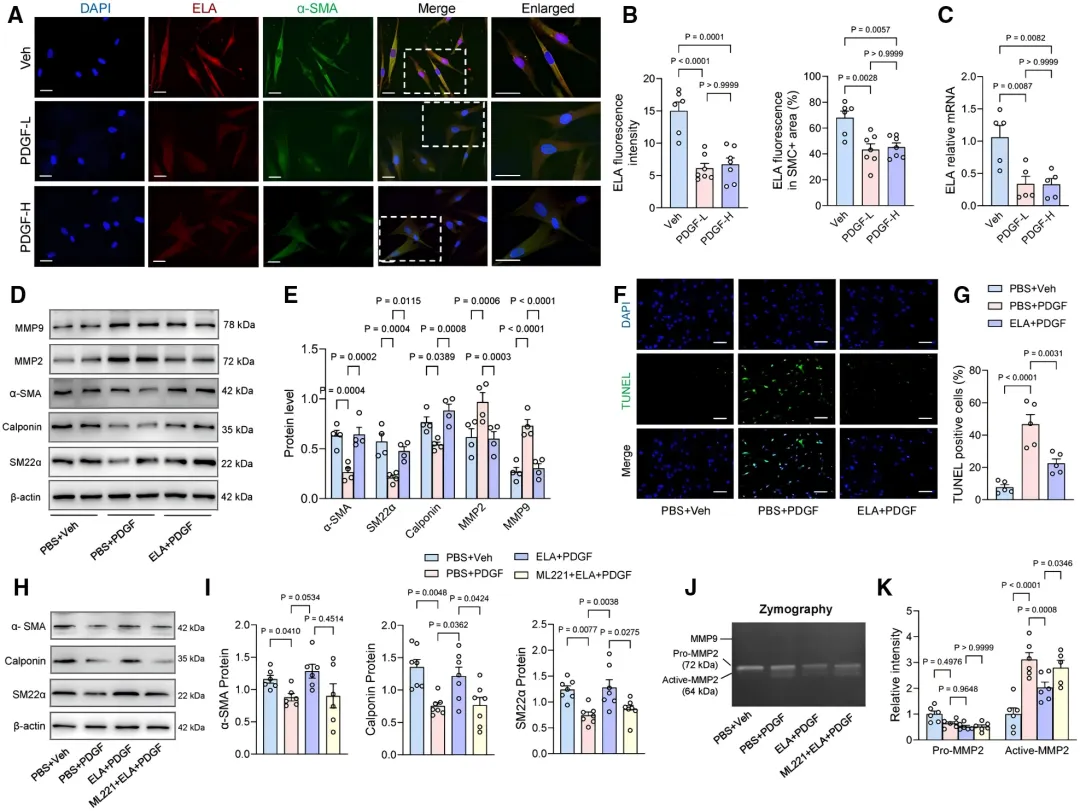
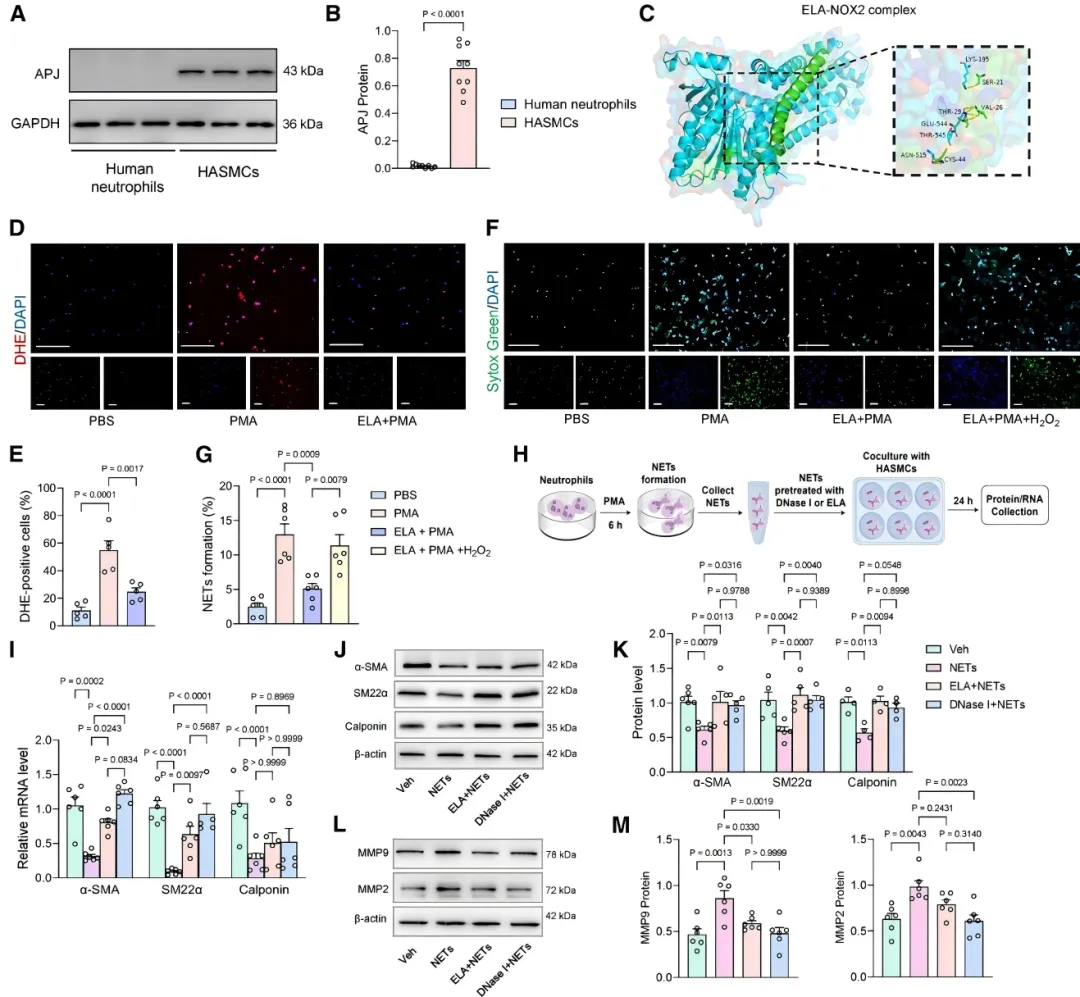
为验证ELA的直接作用,研究者用血小板衍生生长因子(PDGF)处理人主动脉VSMC。PDGF刺激后,细胞内源性ELA表达显著下降【图2A-2C】。
图2.外源性ELABELA(ELABELA)
给药通过APJ
(血管紧张素受体AT1相关受体蛋白)途径,
防止血管平滑肌细胞(VSMC)
稳态紊乱和细胞外基质(ECM)降解
补充外源性ELA则有效逆转了VSMC由收缩向合成表型的转换【图2D】,并抑制了高浓度PDGF引发的细胞凋亡【图2F, 2G】。这些保护作用可被APJ拮抗剂ML221阻断【图2H-2K】,证实ELA通过其受体APJ发挥对VSMC的保护功能。
为进一步在体内验证APJ的关键作用,研究者给TAD小鼠输注ELA的同时加入APJ拮抗剂ML221。
图3.艾拉蛋白(ELABELA)
通过APJ(血管紧张素受体AT1相关受体蛋白)
发挥作用,减轻小鼠胸主动脉夹层(TAD)的病情发展
结果显示,ML221几乎完全逆转了ELA带来的生存获益【图3D】和抑制主动脉扩张的效果【图3E, 3F】。各组小鼠血压和心率无显著差异【图3I-3L】,表明ELA直接作用于血管壁,而非通过影响血流动力学。
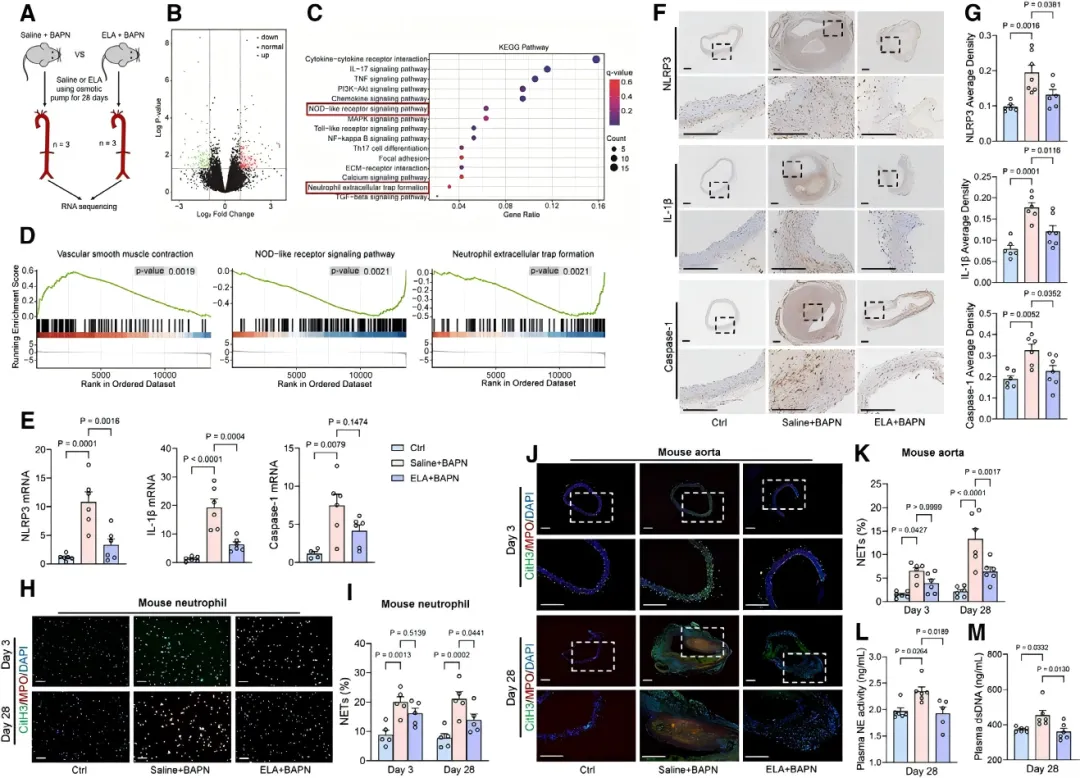
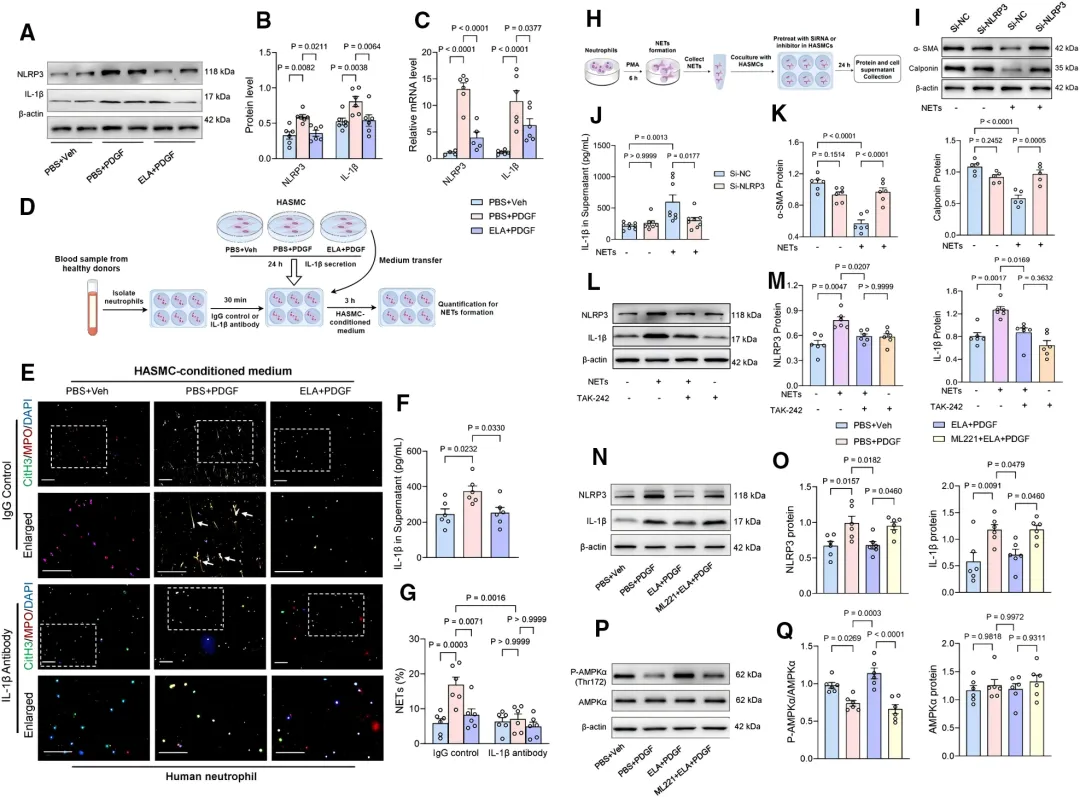
通过主动脉转录组测序分析,研究者发现ELA输注显著抑制了NOD样受体信号通路及NETs形成相关基因的表达【图4C, 4D】。
图4.NLRP3(NLR家族含pyrin结构域蛋白3)
和NET(中性粒细胞胞外诱捕网)
的形成可能在ELABELA(ELA)
对胸主动脉夹层(TAD)的作用机制中发挥作用
进一步验证显示,ELA抑制了主动脉组织中NLRP3炎症小体及IL-1β的激活【图4E-4G】,并减少了循环中NETs标志物水平【图4H-4M】,提示ELA通过抗炎和调控NETs发挥作用。
机制上,研究者发现了一个双向调控环路。首先,ELA直接作用于中性粒细胞,通过抑制NOX2介导的活性氧产生阻断NETs形成【图5C-5G】。
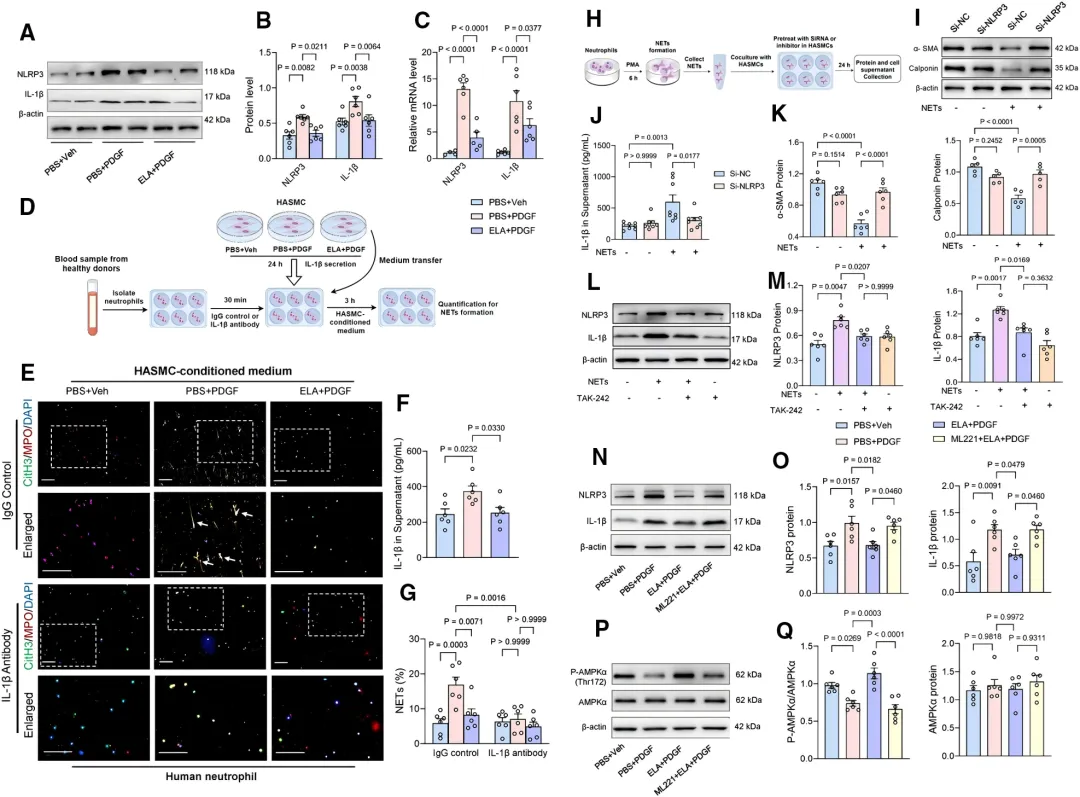
其次,在VSMC中,ELA通过APJ激活AMPK通路,抑制PDGF诱导的NLRP3/IL-1β分泌【图6A-6C, 6P, 6Q】,从而减少由VSMC源性IL-1β触发的NETosis(【图6E, 6G】。
图6.ELA(ELABELA)
通过抑制NLRP3(NLR家族pyrin结构域包含蛋白3)
相关的IL(白细胞介素)-1β产生来
抑制NET(中性粒细胞胞外诱捕网)形成
有趣的是,NETs形成后又会反过来作用于VSMC。
图6.ELA(ELABELA)通过抑制NLRP3
(NLR家族pyrin结构域包含蛋白3)
相关的IL(白细胞介素)-1β产生
来抑制NET(中性粒细胞胞外诱捕网)形成
研究发现,NETs可激活VSMC上的TLR4,触发NLRP3炎症小体活化【图6L, 6M】,导致VSMC自身发生表型转换【图6I-6K】,形成正反馈恶性循环。而ELA能同时作用于循环两端,有效遏制TAD发展。
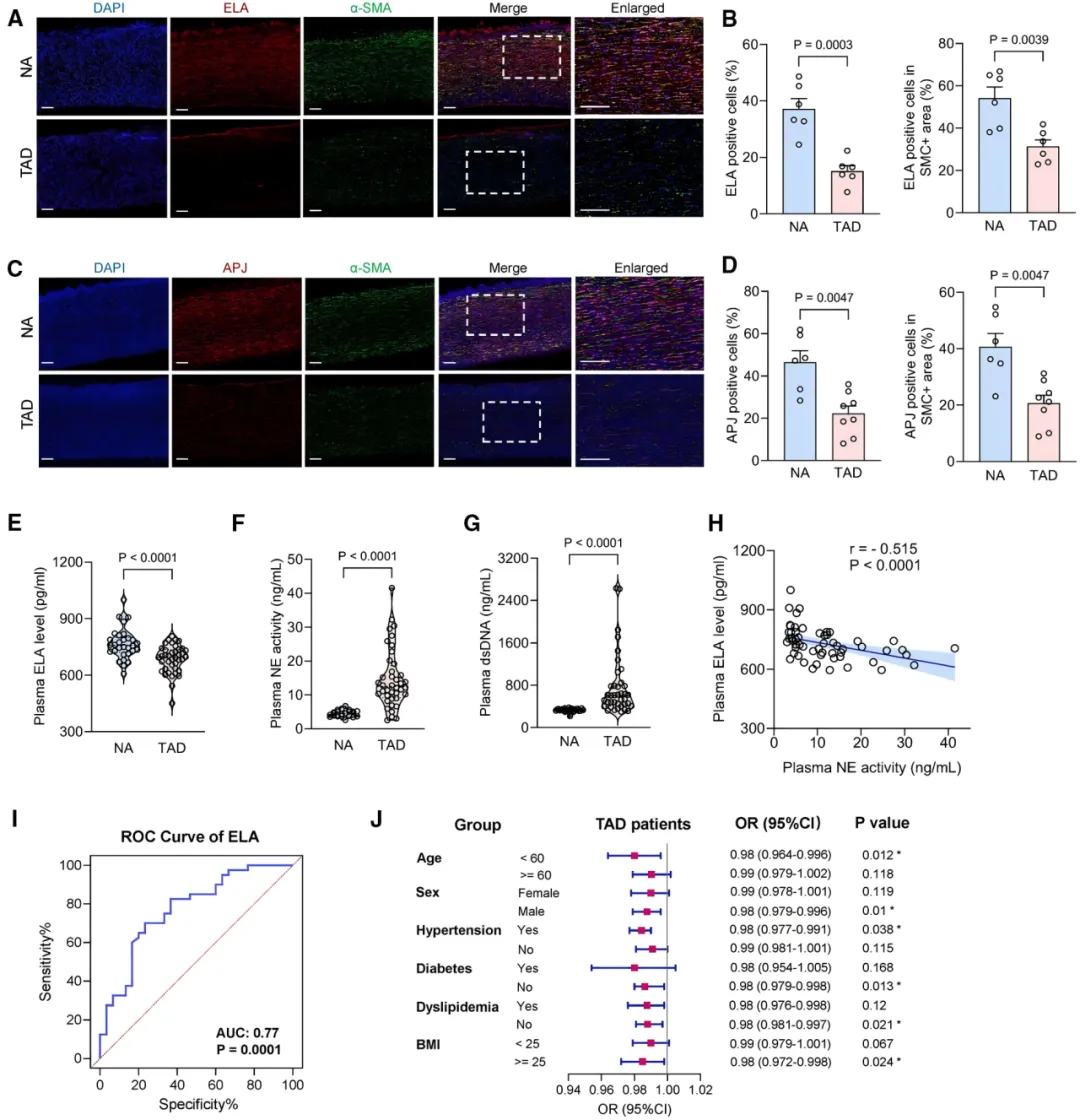
最后,研究者在临床样本中验证了发现的临床意义。与健康对照相比,TAD患者主动脉组织中ELA及APJ受体表达均显著降低【图7A-7D】,血浆ELA水平明显下降而NETs标志物显著升高【图7E-7G】。
图7.ELA(ELABELA)基因表达
在人类胸主动脉夹层(TAD)中呈现下调趋势
相关性分析显示血浆ELA与NETs水平呈负相关【图7H】。低血浆ELA对TAD具有潜在诊断价值【图7I】,且与更高患病风险相关【图7J】,提示ELA有望成为TAD诊断和管理的新型生物标志物。