研究背景与目的
旋毛虫病是严重威胁人类健康的食源性寄生虫病,目前缺乏有效疫苗,主要依赖驱虫药物治疗。核激素受体(NHRs)是寄生虫发育的关键调控因子,其中NHR-23在线虫蜕皮过程中起核心作用,是潜在的药物或疫苗靶点。本研究旨在评估旋毛虫NHR-23的生物学特性及其作为疫苗候选抗原的潜力。具体包括克隆表达TsNHR-23蛋白,分析其在不同发育阶段的转录水平,探究其对肌肉幼虫蜕皮的抑制作用及对巨噬细胞功能的影响,并通过动物实验评估其免疫保护效果。
1 材料与方法
1.1 试验材料
1.1.1 实验动物和虫种 使用SPF级雌性SD大鼠(用于抗体生产及感染血清制备)和ICR小鼠(用于免疫保护试验);旋毛虫虫种为ISSS534株,保存于SD大鼠体内。
1.1.2 菌种、载体和细胞 大肠杆菌DH5α、BL21(DE3);载体pET-32a(+);小鼠巨噬细胞RAW264.7。
1.1.3 主要试剂和材料 各种限制性内切酶、IPTG、弗氏佐剂、PLGA纳米颗粒材料、ELISA试剂盒及各类细胞培养试剂。
1.2 方法
1.2.1 重组质粒pET-32a-TsNHR-23的构建 提取旋毛虫肌肉幼虫RNA,反转录为cDNA,PCR扩增TsNHR-23基因(添加SacⅠ/XhoⅠ酶切位点),连接至pET-32a(+)载体,转化DH5α,经测序验证。
1.2.2 重组TsNHR-23蛋白的表达和纯化 将阳性重组质粒转化BL21(DE3),IPTG诱导表达,镍柱亲和层析纯化蛋白,并去除内毒素。
1.2.3 抗重组TsNHR-23蛋白多克隆抗体的制备 用纯化蛋白乳化弗氏佐剂免疫SD大鼠,多次免疫后采血分离血清。
1.2.4 旋毛虫感染血清的制备 用旋毛虫肌肉幼虫感染大鼠,35 d后采血制备阳性感染血清。
1.2.5 重组蛋白的Western blot分析 检测重组蛋白是否能被旋毛虫感染血清识别(反应原性)。
1.2.6 TsNHR-23基因在旋毛虫不同发育阶段的转录差异 收集成虫、新生幼虫、肌肉幼虫,提取RNA,通过qPCR检测基因转录水平。
1.2.7 抗rTsNHR-23多克隆抗体对肌肉幼虫的影响 体外培养肌肉幼虫,加入不同稀释度的抗rTsNHR-23抗体,显微镜观察并计算蜕皮率。
1.2.8 重组TsNHR23蛋白对小鼠巨噬细胞炎性细胞因子的检测 用不同浓度重组蛋白刺激RAW264.7细胞,qPCR检测IL-1β、IL-6、IL-10等细胞因子的转录水平。
1.2.9 纳米颗粒PLGA包被重组TsNHR-23蛋白 采用双乳化溶剂挥发法制备rTsNHR-23-PLGA纳米颗粒,计算包被效率。
1.2.10 小鼠试验分组与免疫程序 ICR小鼠随机分为PBS组、空载体组、试验组,背部皮下注射免疫(0、14、28 d),第28天进行攻虫感染。
1.2.11 小鼠血清中特异性抗体总IgG的变化检测 采用间接ELISA法检测免疫后不同时间点血清IgG抗体水平。
1.2.12 小鼠血清中细胞因子的检测 ELISA法检测血清中IGF-1、IL-6、TGF-β的变化。
1.2.13 旋毛虫的收集与计数 攻虫后分别收集成虫、新生幼虫和肌肉幼虫,显微镜下计数,计算减虫率。
1.3 数据处理与分析
使用SPSS Statistics 27进行方差分析,GraphPad Prism 10作图。
2 结 果
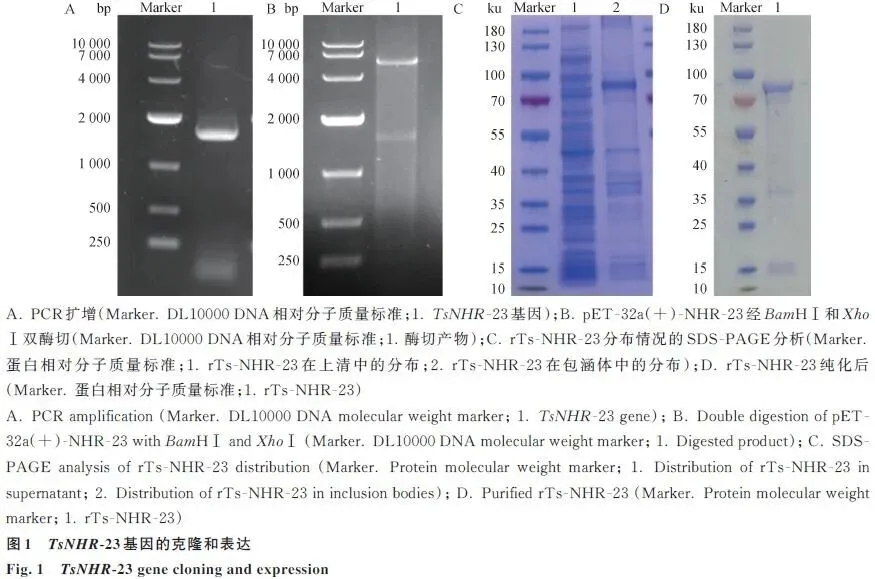
2.1 TsNHR-23基因的克隆和表达
成功扩增出1534 bp的目的基因片段,构建了重组质粒,经酶切鉴定正确。重组蛋白主要以包涵体形式表达,分子量约为70-100 ku,纯化后纯度较高(图1)。
2.2 rTs-NHR-23蛋白的Western blot分析
旋毛虫感染的大鼠血清能在70-100 ku处识别到rTsNHR-23蛋白条带,而阴性血清无反应,证实该蛋白具有良好的免疫反应原性(图2)。

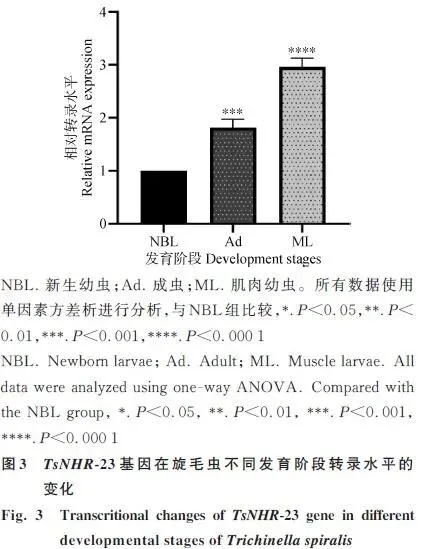
2.3 TsNHR-23基因在旋毛虫不同发育阶段的转录差异
TsNHR-23基因在旋毛虫所有发育阶段均有转录。其中,肌肉幼虫阶段的转录水平最高(为新生幼虫的2.96倍),成虫阶段次之(图3)。
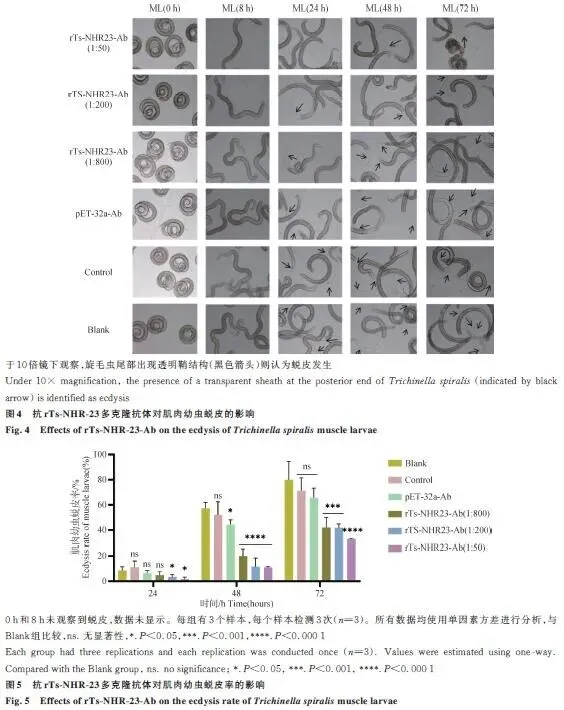
2.4 抗rTs-NHR-23多克隆抗体对肌肉幼虫蜕皮的影响
抗体能显著抑制肌肉幼虫的蜕皮。在48和72 h时间点,不同稀释度的抗体组均表现出极显著的抑制作用(P<0.000 1),且能延缓蜕皮进程(图4、5)。
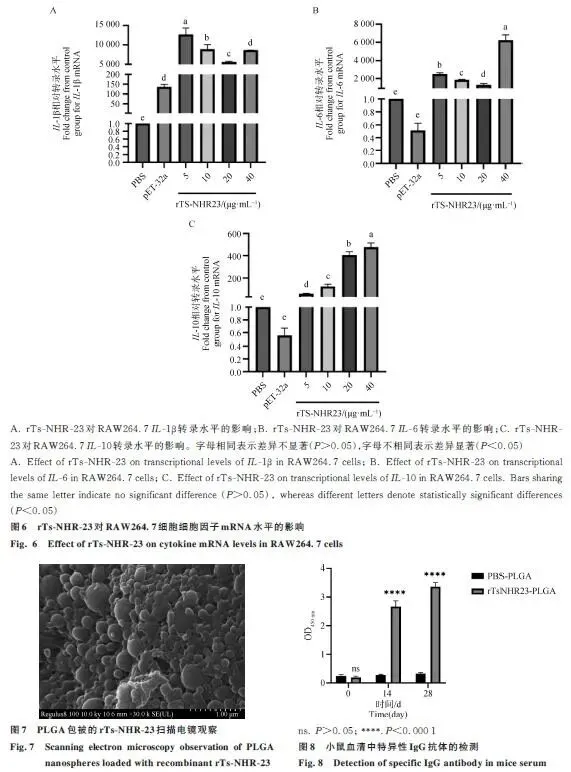
2.5 rTs-NHR-23对小鼠巨噬细胞炎性细胞因子分泌的影响
与对照组相比,不同浓度的rTsNHR-23蛋白均能显著提高RAW264.7细胞中IL-1β、IL-6和IL-10的转录水平,表明其能激活巨噬细胞的炎症反应(图6)。
2.6 PLGA纳米颗粒的包被及包被率的计算
制备的PLGA纳米颗粒呈圆球形。rTs-NHR-23-PLGA的包被效率为72.1%,pET-32a-PLGA为77.6%(图7)。
2.7 小鼠血清中特异性抗体总IgG的变化检测
免疫后,试验组小鼠血清IgG抗体水平显著升高。二次免疫后,试验组抗体滴度维持在高水平,显著高于对照组(P<0.000 1),表明产生了良好的免疫记忆(图8)。
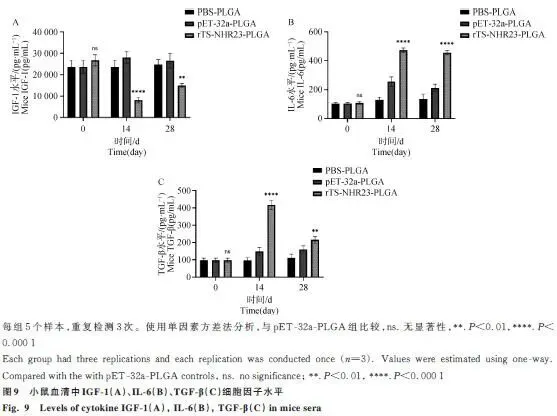
2.8 小鼠血清中细胞因子的检测
IGF-1:首次免疫后显著下降,二次免疫后部分回升但仍低于对照组。IL-6:免疫后显著高于对照组,显示促炎反应。TGF-β:首次免疫后升高,二次免疫后显著下调(图9)。
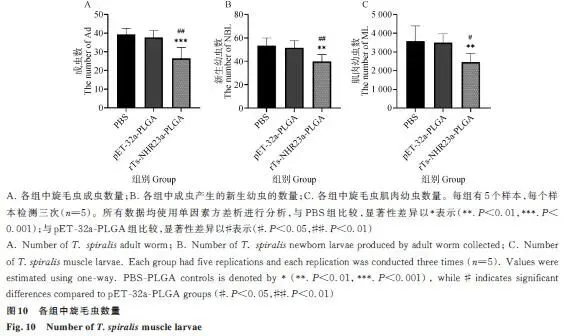
2.9 旋毛虫的收集与计数
成虫减虫率:试验组较空白组降低32.43%。新生幼虫:试验组每条雌虫产幼虫数较空白组下降约25.5%。肌肉幼虫:试验组较空白组减少31.34%(P<0.01)(图10)。表明重组蛋白能显著减少各阶段虫荷。
3 讨 论
免疫原性与功能:rTsNHR-23保留了天然抗原表位,能在肌肉幼虫阶段高表达,且抗血清能抑制幼虫蜕皮,证明其在寄生虫发育中起关键作用。
免疫调节机制:该蛋白能激活巨噬细胞,上调促炎因子(IL-1β, IL-6)和抗炎因子(IL-10),维持免疫平衡。PLGA纳米颗粒有效增强了免疫效果。
免疫应答与保护力:免疫小鼠产生了高水平的IgG抗体和Th1/Th2型细胞因子反应(涉及IGF-1下调和IL-6/TGF-β变化),最终在攻虫试验中显示出显著的减虫效果,证明其作为候选疫苗的潜力。
4 结 论
本研究证实旋毛虫NHR-23在各发育阶段均有分布,其重组蛋白能被宿主免疫系统识别。该蛋白能显著抑制肌肉幼虫蜕皮,并调节巨噬细胞功能。PLGA包被的重组蛋白免疫小鼠后能诱导强烈的体液免疫反应,显著降低旋毛虫的成虫和幼虫负荷,具有作为疫苗候选抗原的开发价值。
关键词:旋毛虫 ; 核激素受体 ; 巨噬细胞 ; 免疫保护