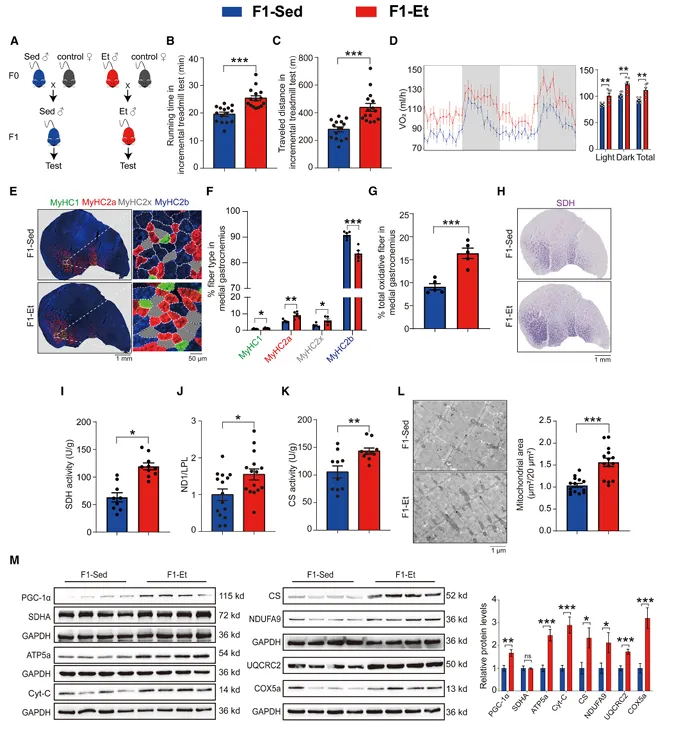
肌肉特异性PGC-1α过表达直接赋予其转基因阴性F1子代高耐力运动能力
为验证PGC-1α在跨代遗传中的作用,研究者使用了MCK-PGC-1α转基因小鼠,其骨骼肌特异性过表达PGC-1α,模拟运动训练的表型。将F0-PGC-1αTG与野生型雌鼠交配,产生转基因阳性和转基因阴性子代(图2)。
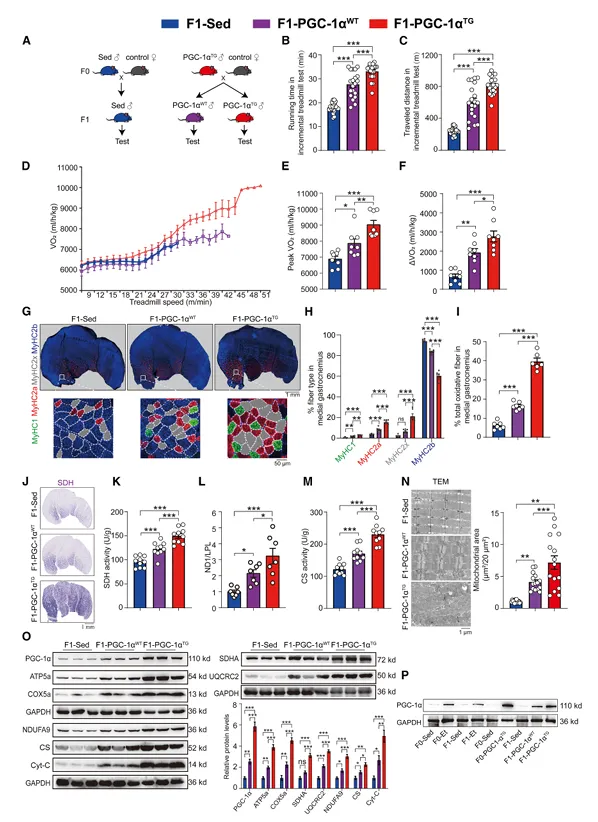
图2. F0肌肉特异性PGC-1α过表达增强其转基因阴性F1子代的耐力运动能
尽管F1-PGC-1αWT不携带转基因,但其耐力运动能力、峰值VO2和ΔVO2均显著优于F1-Sed对照。F1-PGC-1αWT的腓肠肌中氧化型纤维比例、SDH活性、mtDNA含量、CS活性和线粒体蛋白表达均显著高于F1-Sed。
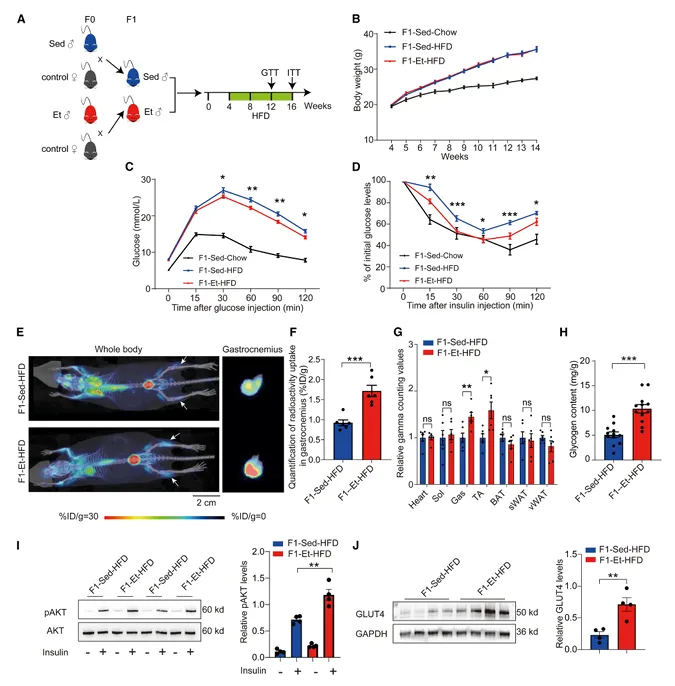
将F1-Et和F1-Sed雄性子代进行12周高脂饮食喂养后,虽然两组体重增加无差异,但F1-Et-HFD的葡萄糖耐量和胰岛素敏感性均显著优于F1-Sed-HFD(图3)。
图3. 父代运动改善F1子代高脂饮食诱导的糖代谢紊乱
18F-FDG PET/CT显像显示,F1-Et-HFD的腓肠肌和胫骨前肌中葡萄糖摄取显著增加,而心脏、比目鱼肌、棕色脂肪和白色脂肪组织无差异。F1-Et-HFD的腓肠肌中糖原含量、AKT磷酸化和GLUT4蛋白表达均显著升高。
精子小RNA(而非长链RNA)介导运动表型的跨代遗传
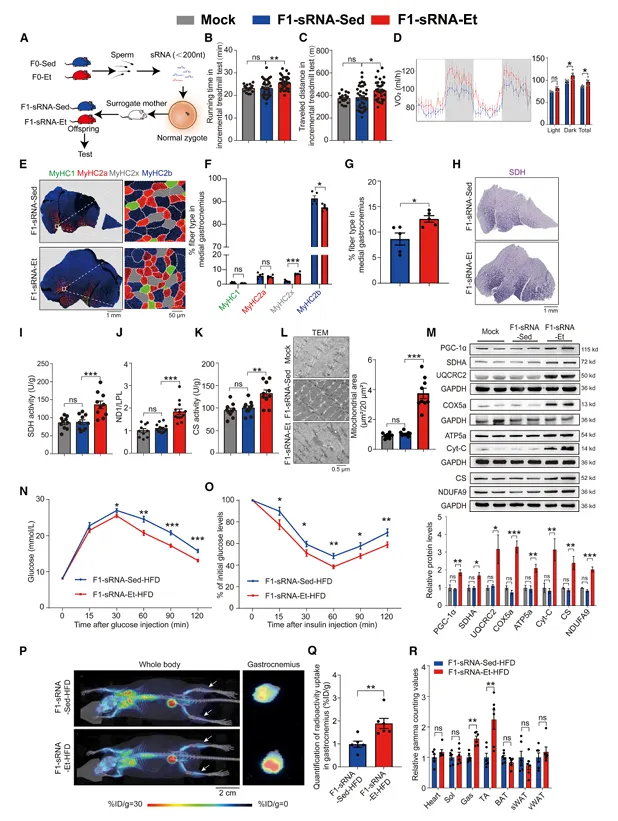
提取F0-Et和F0-Sed精子总RNA,显微注射至正常受精卵后移植,产生的F1子代中,F1-RNA-Et表现出与F1-Et相似的表型:更高的去脂体重、更低的脂肪含量、更强的耐力运动能力、更高的VO2和能量消耗、腓肠肌中更高的氧化型纤维比例、线粒体质量和线粒体蛋白表达(图4)。
图4. 精子小RNA介导父代运动诱导的生理和代谢适应向F1子代的传递
而注射长链RNA的F1子代无任何表型差异。进一步注射精子小RNA的F1子代同样完全复现了运动表型,且F1-sRNA-Et-HFD同样表现出改善的糖耐量和胰岛素敏感性。这些结果证明,精子中的小RNA是介导父代运动获益跨代遗传的关键分子。
运动诱导的精子miRNAs显著改变、转移至卵母细胞并抑制早期胚胎NCoR1表达
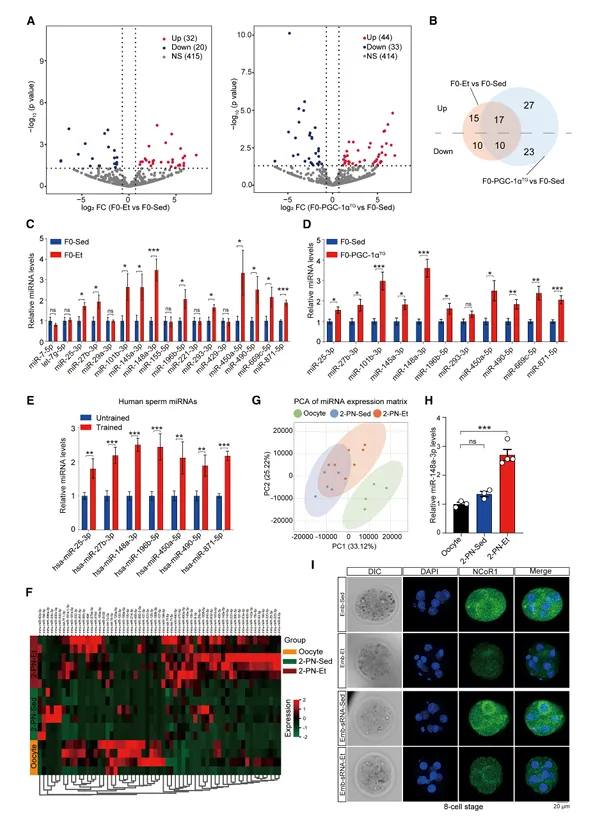
小RNA深度测序显示,F0-Et和F0-PGC-1αTG精子中miRNAs均显著上调,与F0-Sed相比分别鉴定出52个和77个差异表达miRNAs,其中27个miRNAs在两组中共同差异表达(图5)。
图5. 父代运动诱导的精子miRNA变化及其向卵母细胞的转移和对胚胎NCoR1的抑制
在人类中,7个保守miRNAs在耐力训练男性的精子中也显著上调。定量分析显示,运动组精子中miR-148a-3p拷贝数约为静坐组的3倍。对2-细胞期受精卵的测序和RT-qPCR证实,运动来源的精子miRNAs可高效转移至受精卵。GO分析显示miRNAs靶基因富集于“转录辅调节活性”,其中NCoR1是核心靶点。
显微注射miR-148a-3p至正常受精卵可赋予子代高耐力运动能力
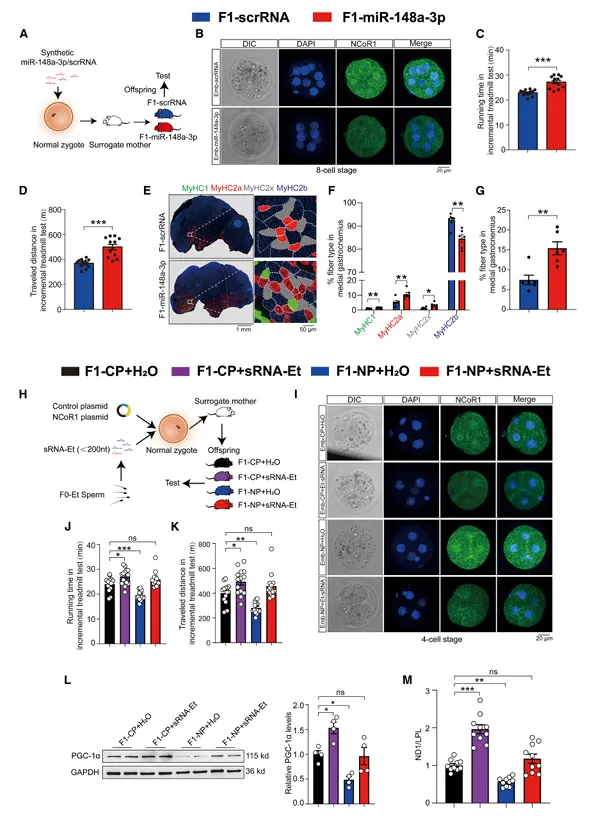
向正常受精卵显微注射生理浓度的合成miR-148a-3p模拟物,产生的F1子代完全复现了运动子代的表型:更高的去脂体重、更低的脂肪含量、更强的耐力运动能力、腓肠肌中氧化型纤维比例增加、线粒体质量增加、线粒体蛋白表达升高(图6)。
图6. 向受精卵注射miR-148a-3p增强子代耐力运动能力
8-细胞期胚胎中NCoR1蛋白和mRNA显著降低,PPARs和ERRα升高。这些结果证实,单个miRNA足以介导父代运动对子代耐力表型的跨代遗传效应。