IF 30.9!南京大学院长领衔!“国自然”创新选题登《cell》子刊!父爱运动,子代天生强!
如果你也想利用国自然热点+多组学进行生信分析,快关注我们后台咨询吧!生信百事通,助力每一个科研梦想!
标题:YAP蛋白可抑制TEAD- NF - κB 复合物活性,并抑制透明细胞肾细胞癌的生长
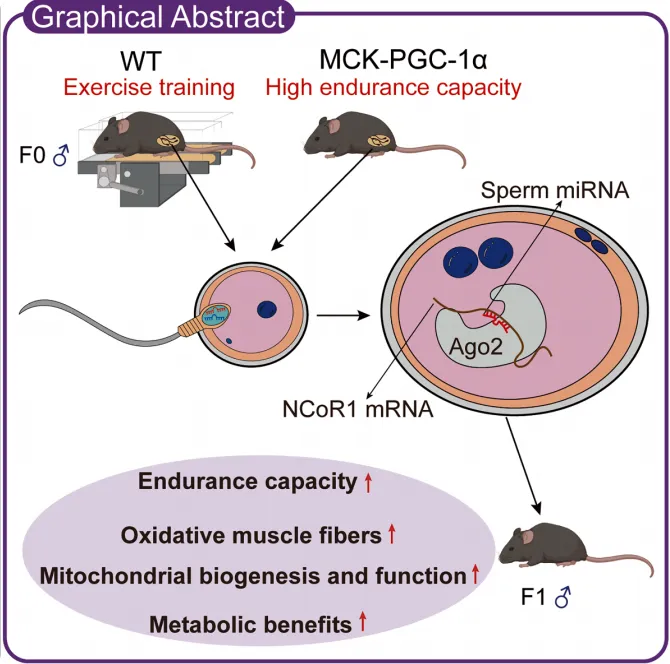
传统观点认为,运动耐力及代谢健康的代际传递主要依赖母系效应,父系运动是否能通过精子表观遗传信息影响后代代谢适应性尚不明确。
本研究旨在揭示父系运动能否及如何通过精子小RNA赋予后代耐力优势。
采用小鼠跑台训练模型和肌肉特异性PGC-1α转基因鼠,通过选择性育种、精子小RNA显微注射及胚胎NCoR1过表达挽救实验,结合运动耐力测试、代谢分析和小RNA测序,揭示父系运动代际遗传的关键分子通路。
📌三大维度的研究思路框架已整理,点击图片查看。(5分钟获取核心要点)!
《首建“精子miRNAs-NCoR1”轴:
父系运动跨代遗传新范式》
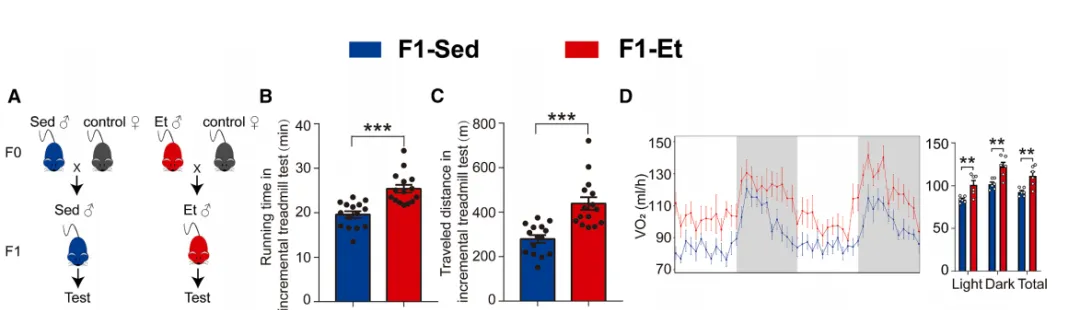
为探究运动获益能否跨代传递,研究者对雄性小鼠进行8周 treadmill 训练(F0-Et),随后与正常雌鼠交配产生子代(F1-Et)【图1A】。
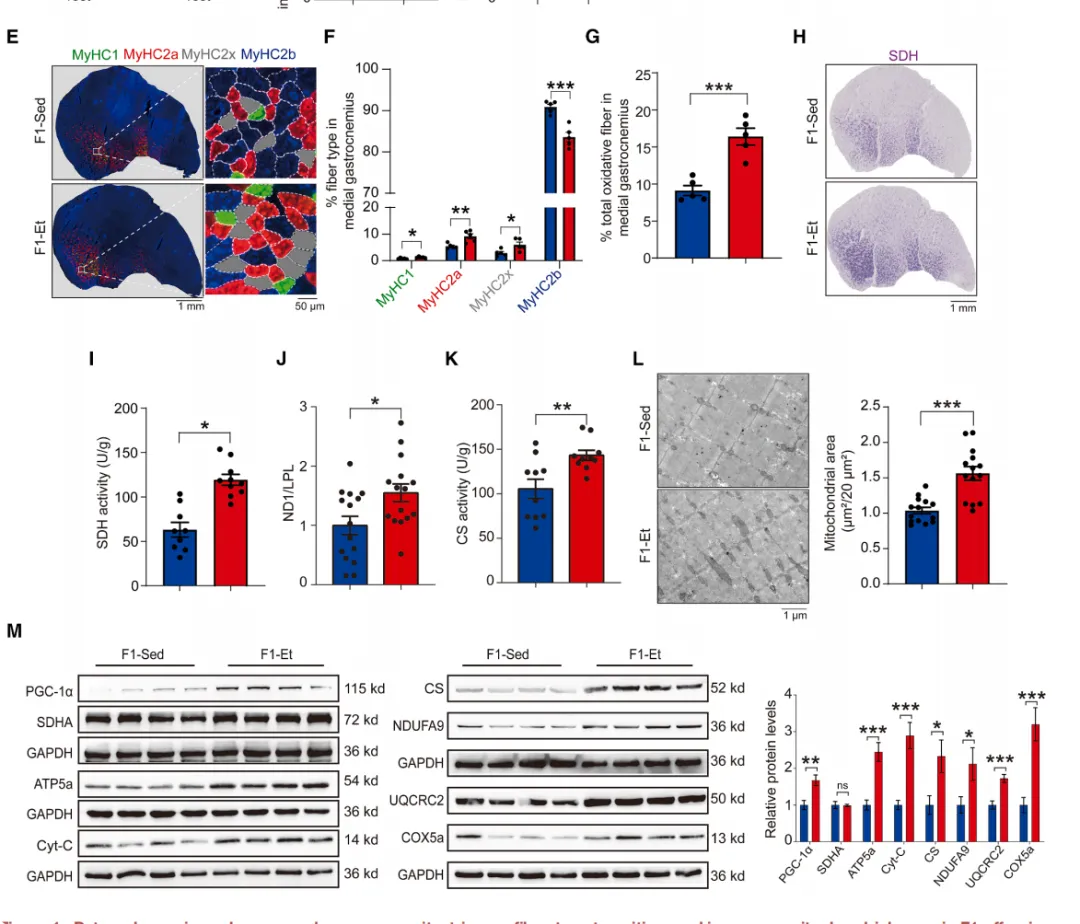
图1.父系运动可增强F1代子代的耐力能力、
触发肌纤维类型转换并增加线粒体质量
结果令人振奋:F1-Et子代跑动距离和持续时间均显著超过 sedentary 父系子代(F1-Sed)【图1B-C】,耗氧量和能量消耗也更高【图1D】,而心脏功能无差异。
耐力提升的背后是肌肉发生了“质的改变”。免疫荧光显示,F1-Et子代 gastrocnemius 中氧化型肌纤维比例显著增加,糖酵解型IIb纤维减少【图1E-G】。
图1.父系运动可增强F1代子代的耐力能力、
触发肌纤维类型转换并增加线粒体质量
SDH染色和电镜证实其线粒体密度更高【图1H-L】,PGC-1α及多种线粒体标志蛋白均显著上调【图1M】。
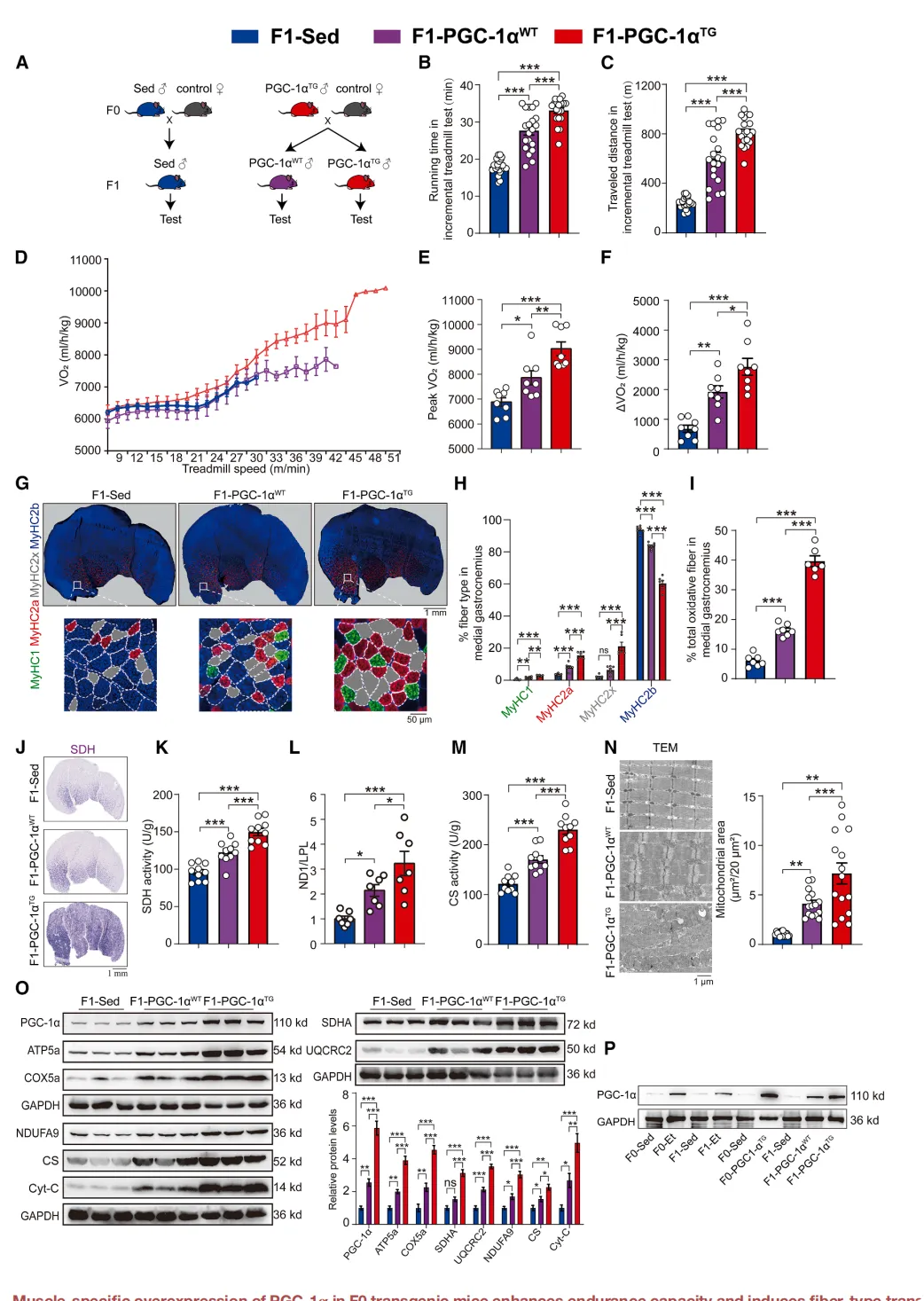
为验证PGC-1α的关键作用,研究者将F0-PGC-1α TG雄鼠与野生型雌鼠交配,获得未继承转基因的野生型子代(F1-PGC-1α WT)【图2A】。
图2.肌肉特异性 PGC-1α 过表达
增强 F1 代耐力并诱导肌纤维转变
及线粒体生物合成
结果令人意外:这些“空转”子代仍表现出更强耐力,峰值VO₂提升14%【图2B-E】,氧化纤维增加,线粒体增多【图2G-N】,表型与F1-Et高度相似。
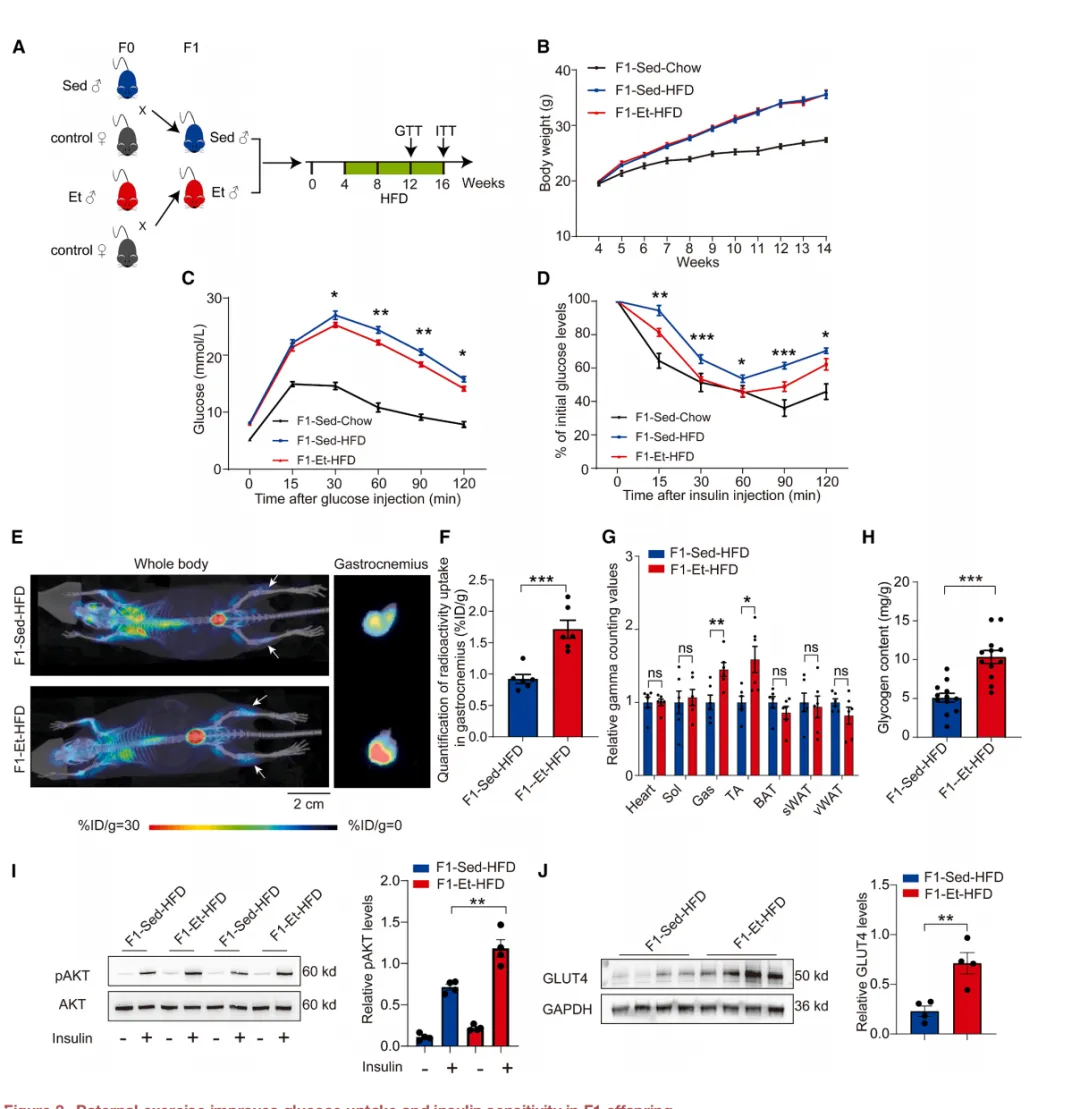
运动遗传的益处不仅限于耐力。研究者对F1子代进行12周高脂饮食喂养【图3A】。
图3.父系运动可改善F1代子代
的葡萄糖摄取及胰岛素敏感性
结果令人鼓舞:F1-Et-HFD组的葡萄糖耐量和胰岛素敏感性均显著优于F1-Sed-HFD组【图3C-D】。
PET/CT显示其骨骼肌葡萄糖摄取量更高【图3E-G】,AKT磷酸化和GLUT4表达显著增强【图3I-J】。
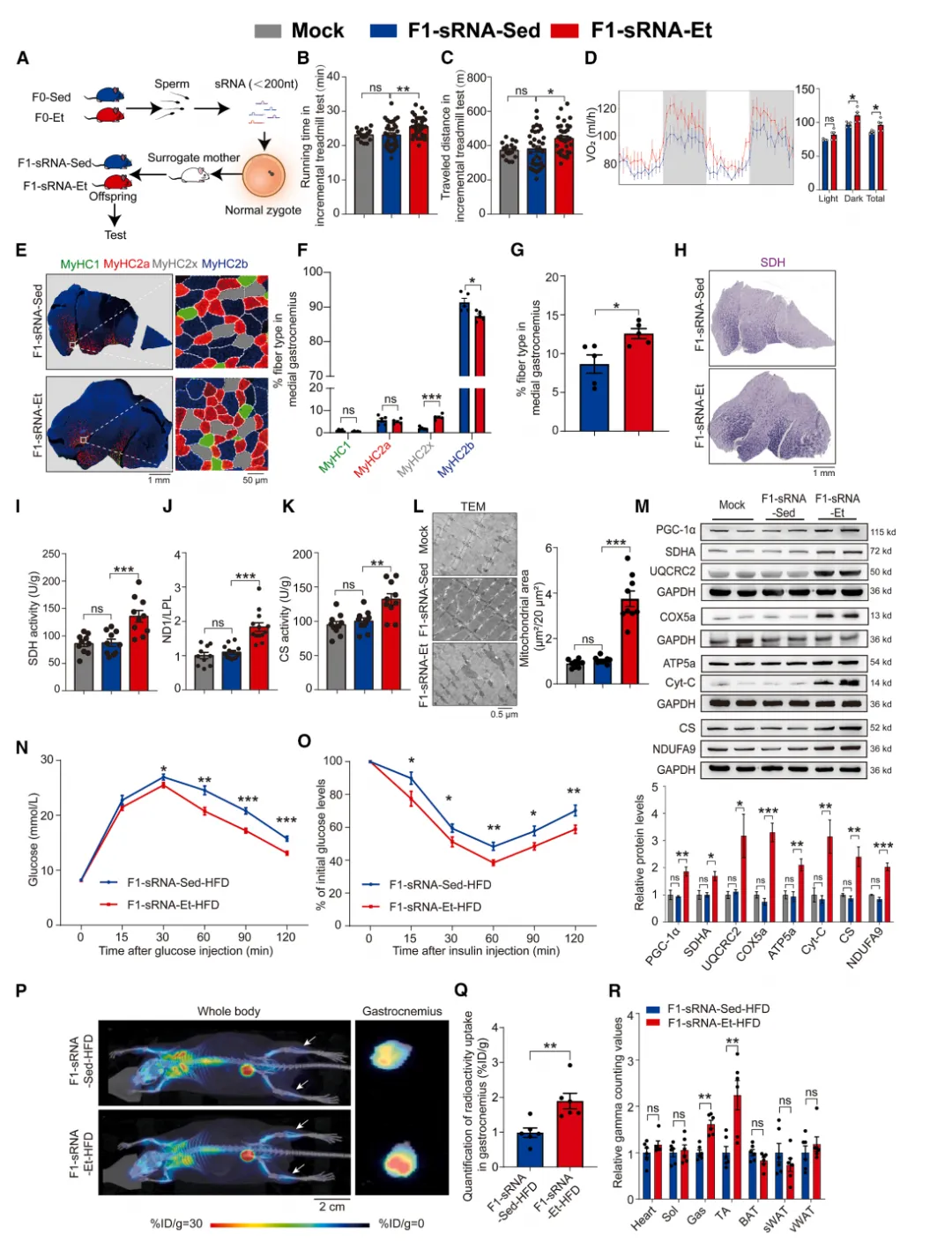
那么,运动信息如何写入精子?研究者提取运动父鼠精子总RNA,显微注射至正常受精卵中。
图4.精子小RNA介导父系运动诱导
的生理及代谢适应性向 F1 代传递
结果极具说服力:仅注射小RNA(sRNA)的子代(F1-sRNA-Et)就完整重现了运动表型——耐力提升、VO₂升高、氧化纤维增加【图4B-G】。高脂喂养实验进一步证实注射sRNA可改善子代葡萄糖稳态【图4N-R】。
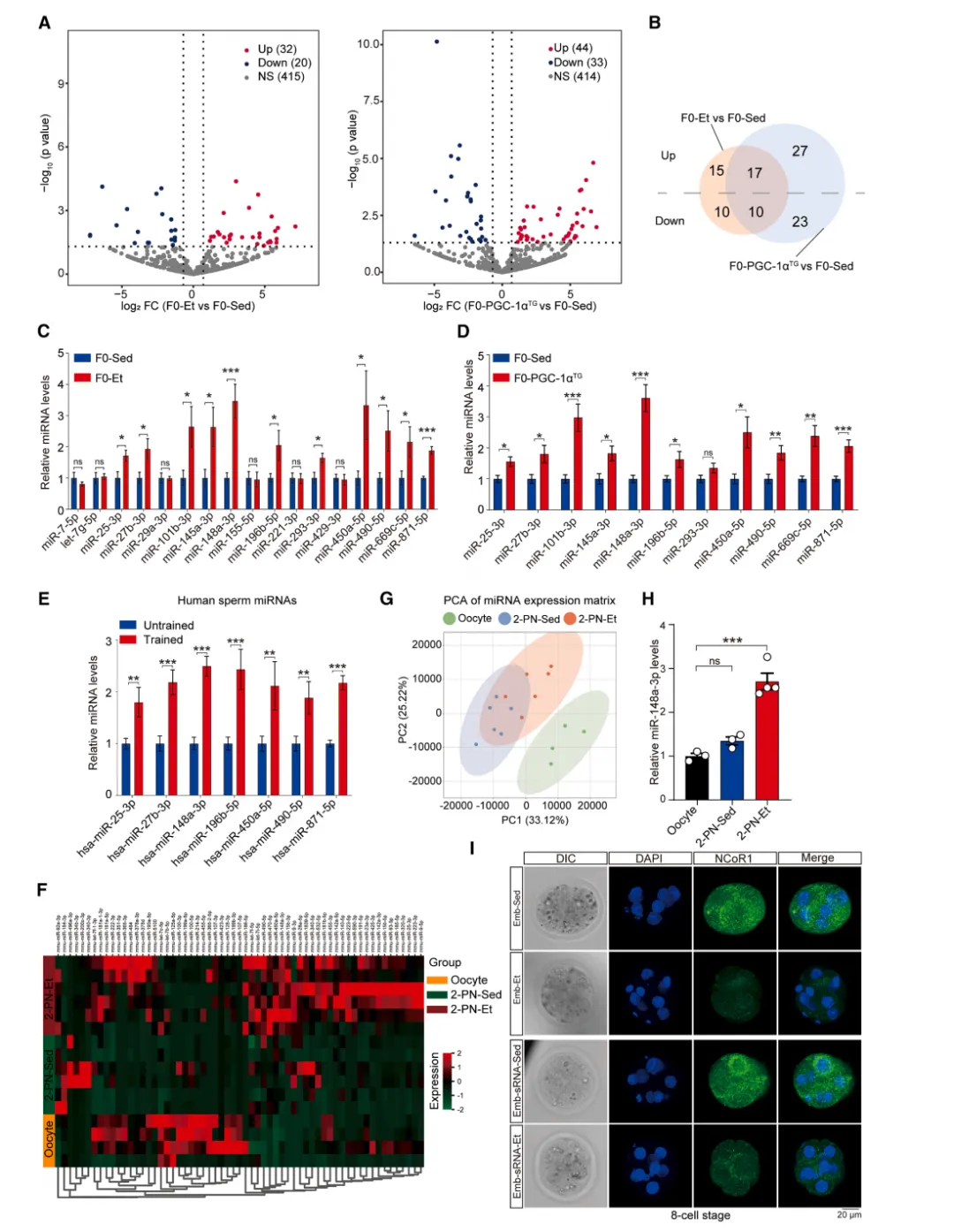
为明确具体分子,研究者对运动父鼠精子进行小RNA测序。结果显示,F0-Et和F0-PGC-1α TG的精子miRNA表达模式高度相似,均与sedentary组显著区分【图5A-B】。
图5.运动训练重塑精子 miRNA
谱并有效传递至卵子
RT-qPCR验证了11个miRNA在运动组中显著上调,其中10个在两模型中一致【图5C-D】。
在人类训练者精子中,7个保守miRNA也显著升高【图5E】。
这些精子miRNA进入胚胎后做了什么?研究者检测了受精后2-原核期胚胎,发现运动父鼠来源的miRNA(如miR-148a-3p)成功传递至胚胎【图5H】。
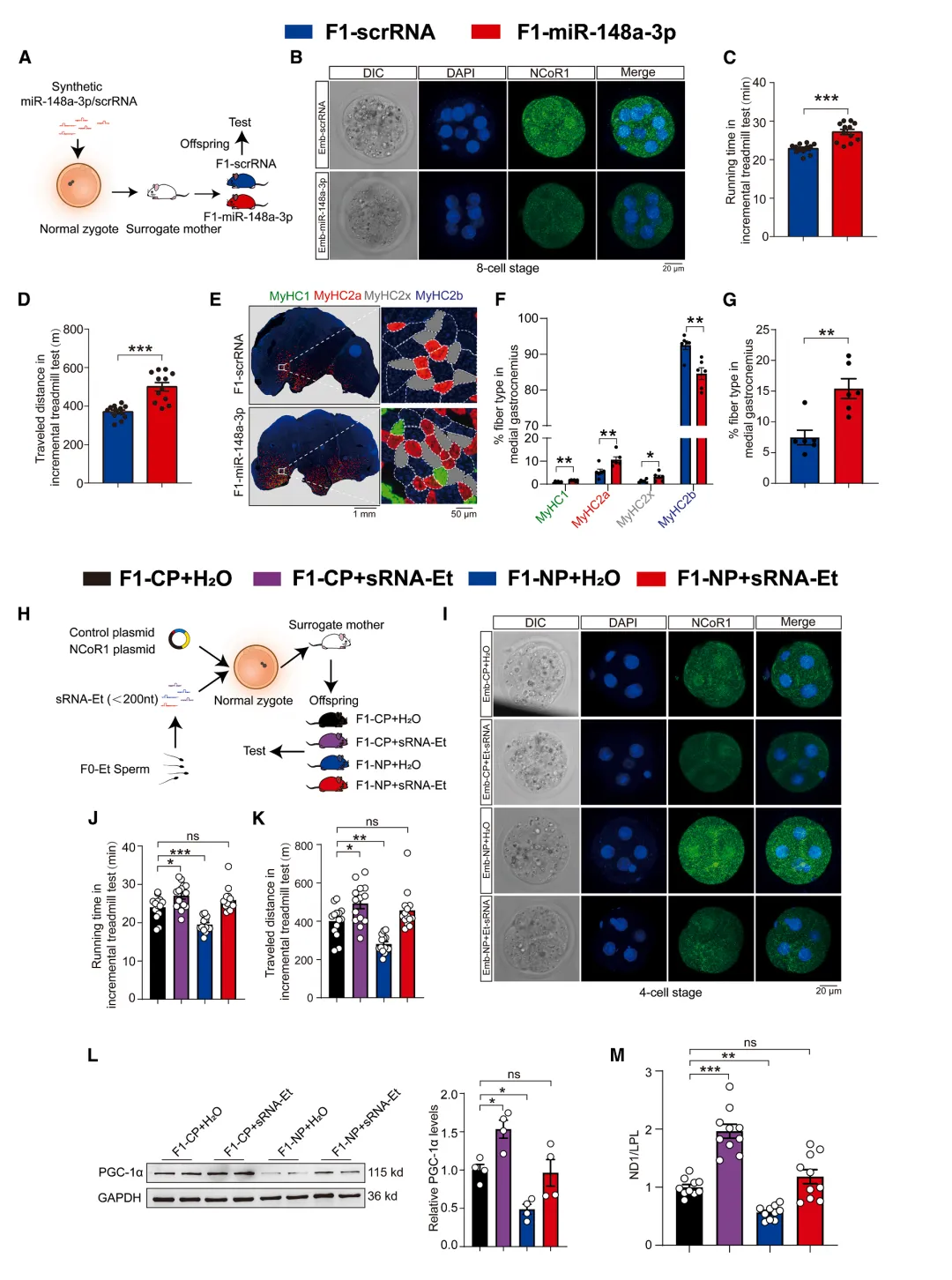
图6.合子注射 miR-148a-3p 增强子代耐力,
早期胚胎恢复 NCoR1 表达可减弱该效应
8细胞期胚胎免疫荧光显示,注射运动父鼠sRNA后,NCoR1蛋白水平显著降低【图5I-J】。
共注射NCoR1过表达质粒可完全抵消sRNA诱导的表型【图6H-M】。
本研究发现,父系运动通过上调精子中的特定miRNA(如miR-148a-3p),在早期胚胎中抑制NCoR1表达,从而重编程代谢网络,促进后代骨骼肌线粒体生成和氧化型肌纤维转化,最终增强耐力并改善糖代谢。
有生信分析需求的同学快来联系生信百事通吧!有消息一定及时回复!























 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
