3.1 ABA负向调节大豆对大豆疫霉的免疫反应
ABA含量降:疫霉菌侵染后大豆内源ABA含量显著下降,合成基因 GmNCED3 表达下调而代谢基因 GmCYP707 表达上调,信号通路基因转录水平发生显著变化。
促病害加重:外源施加ABA或其抑制剂氟啶酮对病原菌体外生长无直接影响,但在植物体内ABA显著扩大了病斑面积并增加了病原菌生物量,而氟啶酮则减轻了发病症状。
抑抗病基因:转录组测序分析表明,ABA处理导致几丁质响应、防御反应等相关基因显著下调,特别是NBS-LRR、WRKY和PR等抗病基因家族的表达受到显著抑制。
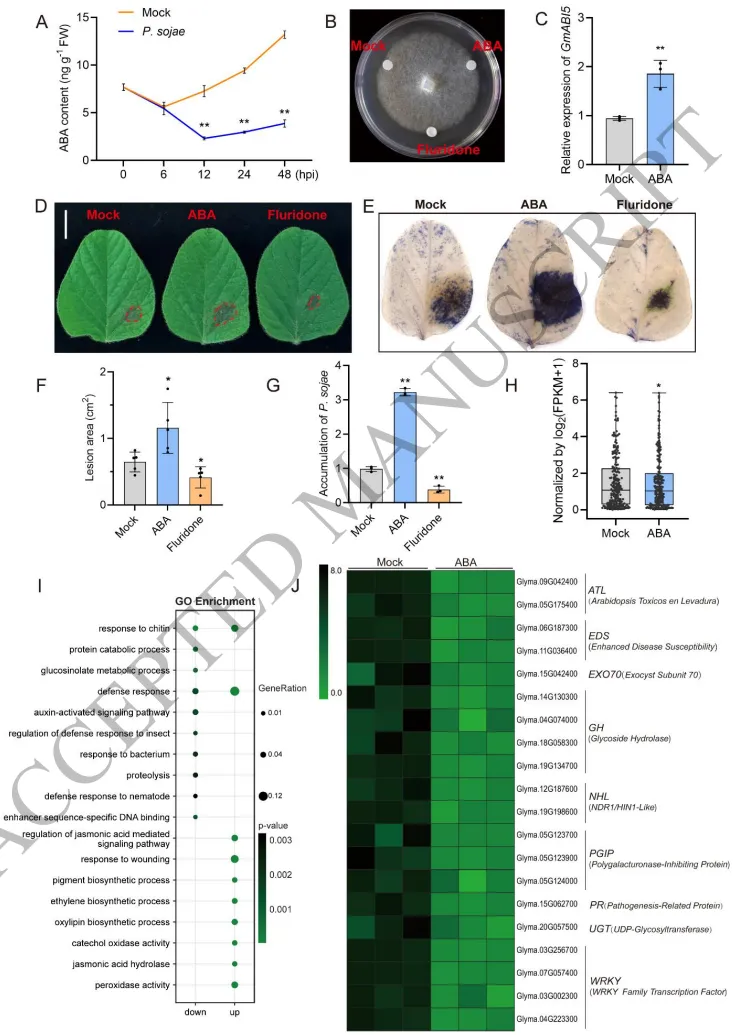
大豆在受到大豆疫霉 (P. sojae) 侵染后,其体内内源脱落酸 (ABA) 含量显著下降,且伴随着ABA合成基因的下调与代谢基因的上调。体外试验表明ABA及其抑制剂氟啶酮并不直接影响病原菌的生长,但在大豆植株上,外源ABA处理显著加重了病害症状并增加了病原菌生物量,而氟啶酮则减轻了病害。转录组分析进一步揭示,ABA处理广泛抑制了植物的防御反应路径,导致大量抗病相关基因家族(如NBS-LRR、WRKY和PR)的表达显著下调,从而削弱了宿主的免疫防御能力 (Fig. 1)。
图1. 脱落酸负向调节大豆免疫。 A) 接种大豆疫霉 (P. sojae) 的大豆叶片中内源脱落酸 (ABA) 含量。B) P. sojae 在添加ABA或氟啶酮的V8培养基上的菌丝生长情况。C) P. sojae 侵染下 GmABI5 的相对表达水平。GmActin11 用作内参。D) 接种 P. sojae 的大豆叶片表型。比例尺 = 1 cm。E) 接种 P. sojae 的叶片的台盼蓝染色。F) 图 (D) 中所示病斑面积的定量分析。G) 受侵染大豆叶片中 P. sojae 的相对生物量。计算 P. sojae cDNA (PsActin) 与大豆cDNA (GmActin11) 的比值,并以Mock对照(设为1)进行归一化。H) NBS-LRR(核苷酸结合位点富含亮氨酸重复序列)基因家族的转录水平,通过 log₂(FPKM+1) 进行归一化。箱线图元素:中线,中位数;箱体边界,上下四分位数;须线,1.5 × 四分位距;点,异常值。I) 差异表达基因 (DEGs) 的基因本体论 (GO) 分析。J) 展示抗病相关基因家族转录水平的热图。彩色方块表示所选基因的转录值,从0(绿色)到8(黑色),通过 log₂(FPKM+1) 进行归一化。转录组数据来源于“P. sojae 侵染下经ABA处理的大豆叶片(来自10日龄幼苗)” (H, I, J)。柱状图数值代表平均值 ± SD (A, C 和 G: n = 3; F: n = 5; H: n = 210)。B、D 和 E 中的试验独立重复三次,结果相似。星号表示与Mock相比具有统计学显著差异 (Student's t检验):*, P < 0.05; **, P < 0.01。
3.2 GmABI5抑制大豆免疫反应从而降低对大豆疫霉的抗性
正调控感病:疫霉菌侵染诱导了 GmABI5 的转录,过表达 GmABI5 的大豆毛状根中病原菌积累显著增加且菌丝扩展加快,而敲低该基因则显著降低了病原菌的积累与扩展速率。
抑防御网络:对过表达转基因植株的转录组分析显示,无论在正常生长还是病原菌侵染条件下,大量与防御反应、过氧化物酶活性及细胞壁合成相关的基因均被显著下调。
抗性基因降:进一步分析表明,GmABI5 的过表达导致NBS-LRR等核心抗病基因家族的表达水平显著降低,全面抑制了植物的免疫反应。
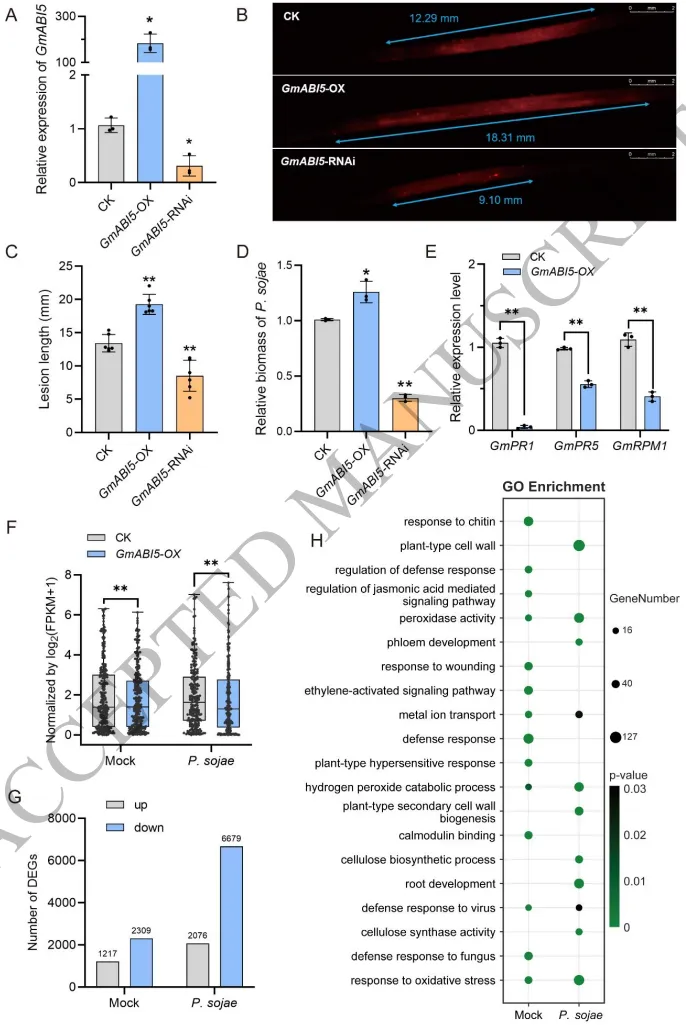
大豆疫霉 (P. sojae) 侵染能够诱导依赖于ABA的 GmABI5 转录表达。通过构建转基因大豆毛状根发现,过表达 GmABI5 会显著促进病原菌的积累和菌丝扩展,加重病害症状;相反,敲低该基因则能有效减缓病菌侵染,提升植物抗性。转录组学研究进一步阐明,GmABI5 的过表达无论在正常条件还是侵染状态下,均广泛抑制了与几丁质响应、活性氧清除以及纤维素合成相关的防御通路,并显著下调了NBS-LRR等核心抗病基因家族的表达,从而在转录水平上削弱了大豆的免疫防御网络 (Fig. 2)。
图2. GmABI5负向调节大豆对大豆疫霉的抗性。 A) 毛状根中 GmABI5 的相对表达水平。GmActin11 基因用作内参。B) 接种RFP标记的大豆疫霉 (P. sojae) 的大豆毛状根的代表性表型。图像在接种后24小时 (hpi) 使用荧光体视显微镜拍摄。比例尺 = 2 mm。C) 图 (B) 中所示病斑长度的定量分析。D) P. sojae 的相对生物量。PsActin 用作 P. sojae 的内参。GmActin11 用作大豆的内参。相对病原菌生物量计算为 P. sojae cDNA 与大豆cDNA的比值,对照 (CK) 组的比值设为1。E) GmPR1(发病相关蛋白1)、GmPR5(发病相关蛋白5)和 GmRPM1(抗病蛋白1)的相对表达水平。GmActin11 用作内参。F) NBS-LRR(核苷酸结合位点富含亮氨酸重复序列)基因家族的转录水平,通过 log₂(FPKM+1) 进行归一化。G) 与对照 (CK) 相比,GmABI5-OX毛状根中上调和下调的DEGs数量。H) 下调的差异表达基因 (DEGs) 的基因本体论 (GO) 分析。转录组数据来源于“P. sojae 侵染下的 GmABI5-OX 和 CK 毛状根” (F, G, H)。柱状图数值代表平均值 ± SD (A, D 和 E: n = 3; C: n = 6; F: n = 230)。B 中的试验独立重复三次,结果相似。星号表示与对照 (CK) 相比具有统计学显著差异 (Student's t检验):*, P < 0.05; **, P < 0.01。
3.3 GmABI5与GmCDK8发生物理相互作用
核内共定位:亚细胞定位分析表明GmABI5蛋白定位于细胞核中,为探究其调控机制,通过核系统酵母双杂交筛选鉴定出互作蛋白GmCDK8。
体外互作证:体外Pull-down试验证实,带有GST标签的GmCDK8能够成功拉下带有MBP标签的GmABI5,表明两者在体外具有直接结合能力。
体内互作证:荧光素酶互补成像、双分子荧光互补以及免疫共沉淀试验进一步证实了GmABI5与GmCDK8在植物细胞核内存在真实的物理相互作用。
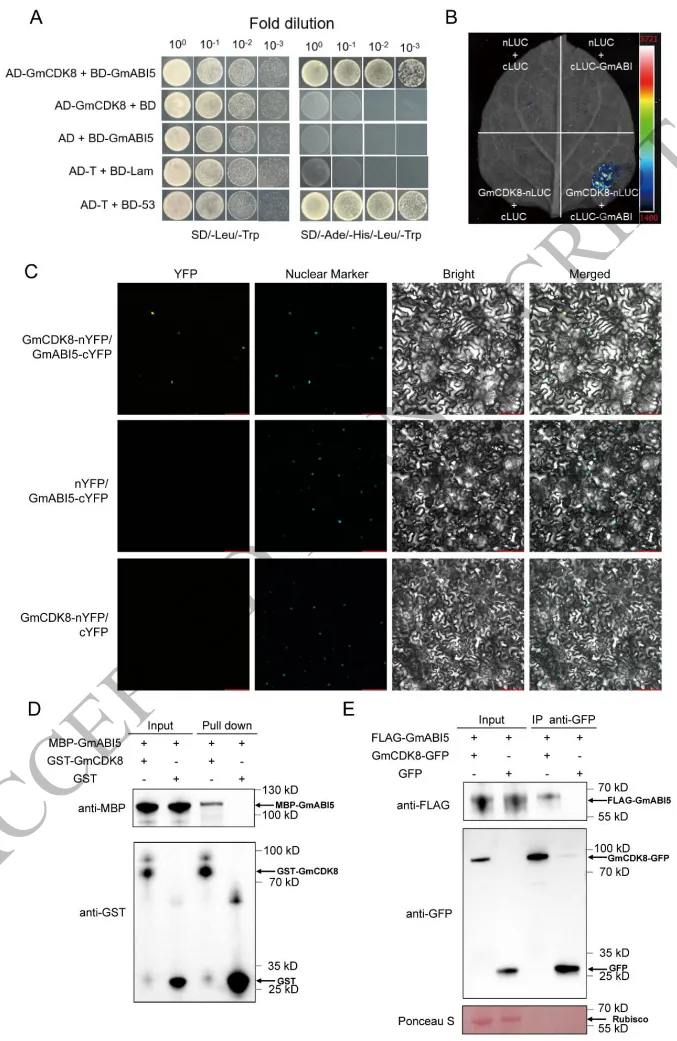
亚细胞定位显示GmABI5定位于细胞核内。为了揭示其免疫调控机制,通过酵母双杂交筛选发现GmCDK8是其潜在的互作蛋白,且体外Pull-down试验证实了两者之间的直接结合。随后,在植物体内进行的荧光素酶互补成像 (LCI) 和双分子荧光互补 (BiFC) 试验均检测到了特异性的核内互作信号。此外,免疫共沉淀 (Co-IP) 试验也成功验证了这一结合关系。这些体内外试验结果充分证明了GmABI5与GmCDK8之间存在直接的物理相互作用 (Fig. 3)。
图3. GmABI5与GmCDK8发生物理相互作用。 A) 酵母双杂交 (Y2H) 试验显示GmABI5与GmCDK8之间的相互作用。B) 荧光素酶互补成像 (LCI) 试验确认了在本氏烟草 (N. benthamiana) 叶片中的相互作用。C) 双分子荧光互补 (BiFC) 试验显示在本氏烟草叶表皮细胞中的相互作用。比例尺 = 100 μm。D) Pull-down试验显示带有GST标签的GmCDK8在体外结合带有MBP标签的GmABI5。E) 免疫共沉淀 (Co-IP) 试验显示GmABI5与GmCDK8之间的相互作用。带有FLAG标签的GmABI5与固定在GFP磁珠上的带有GFP标签的GmCDK8蛋白共同孵育。
3.4 GmCDK8负向调节大豆对大豆疫霉的抗性
受多信号诱导:GmCDK8定位于细胞核和细胞膜,其转录水平受到flg22、水杨酸、茉莉酸和脱落酸等多种免疫和激素信号的显著诱导。
负调控抗性:接种试验表明,gmcdk8 突变体表现出病斑减小和菌丝扩展受抑的抗病表型,而过表达 GmCDK8 的植株则表现出更为严重的感病症状。
抑抗病基因:分子水平分析显示,在 gmcdk8 突变体中,多种疾病抗性基因的表达水平得到显著上调,进一步印证了其在防御反应中的负调控角色。
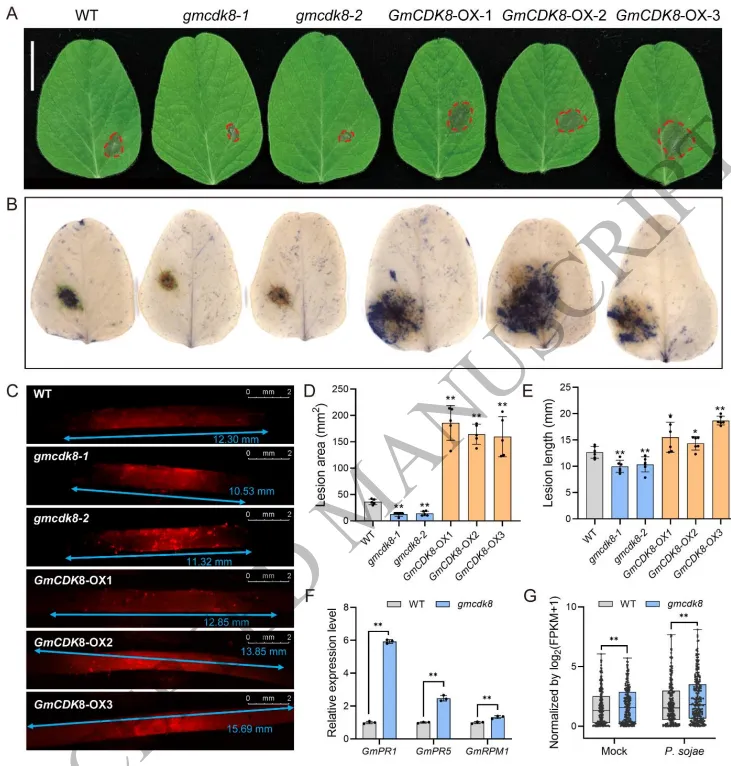
亚细胞定位显示GmCDK8同时存在于细胞核和细胞膜中,且其转录表达可被flg22、水杨酸 (SA)、茉莉酸 (JA) 和脱落酸 (ABA) 等多种信号显著诱导,暗示其参与了免疫调节过程。通过对突变体和过表达植株的表型分析发现,gmcdk8 突变体在接种大豆疫霉后病斑面积减小、菌丝扩展受阻,表现出增强的抗病性;而 GmCDK8 过表达植株则表现出更高的病原菌生物量积累和更严重的坏死病斑。此外,抗病相关基因在 gmcdk8 突变体中显著上调。这些结果一致表明,GmCDK8在大豆抵抗大豆疫霉的过程中发挥着负向调节作用 (Fig. 4)。
图4. GmCDK8负向调节大豆对大豆疫霉的抗性。 A, B, D) 接种大豆疫霉 (P. sojae) 游动孢子后,野生型 (WT)、GmCDK8 过表达株系 (GmCDK8-OX) 和突变体 (gmcdk8) 的病斑表型及病斑面积定量分析。C, E) 接种RFP标记的 P. sojae 的大豆根部菌丝扩展表型及红色荧光菌丝长度的定量分析。F, G) gmcdk8 突变体中疾病抗性相关基因的相对表达水平。
3.5 GmCDK8与GmABI5相互促进形成正反馈回路
促蛋白稳定:无细胞蛋白降解和共表达试验表明,GmCDK8能够通过部分抑制泛素-蛋白酶体途径,延缓GmABI5的降解,显著提高其蛋白丰度。
非直接磷酸化:尽管GmCDK8具有激酶活性,但体外磷酸化试验未能观察到其对GmABI5的直接磷酸化修饰,暗示可能存在其他辅助因子或间接机制。
转录正反馈:基因沉默和双荧光素酶报告试验证实,病原菌侵染或flg22处理能触发GmABI5对 GmCDK8 启动子的转录激活,形成相互促进的正反馈回路。
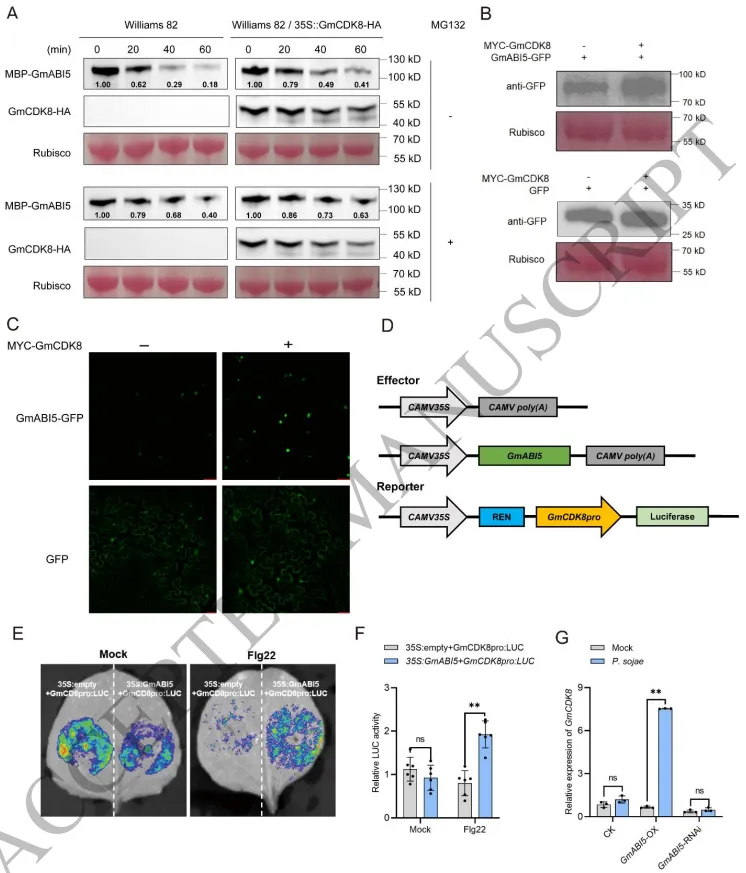
为了探究GmCDK8对GmABI5的调控机制,无细胞蛋白降解和植物体内共表达试验揭示,GmCDK8能够通过部分阻断依赖于26S蛋白酶体的降解途径,有效延缓GmABI5蛋白的降解并提升其积累量。尽管体外试验未能直接检测到GmCDK8对GmABI5的磷酸化作用,但进一步的转录调控分析发现,GmABI5 的沉默会降低 GmCDK8 的表达。更为重要的是,病原菌侵染或flg22处理能够特异性地触发GmABI5对 GmCDK8 启动子的转录激活。这种GmCDK8稳定GmABI5蛋白与GmABI5促进 GmCDK8 转录的相互作用,在植物免疫响应中构建了一个放大信号的正反馈回路 (Fig. 5)。
图5. GmCDK8与GmABI5相互促进形成正反馈回路。 A) 无细胞蛋白降解试验检测GmABI5蛋白的稳定性。在添加或不添加蛋白酶体抑制剂MG132的情况下,将重组GmABI5蛋白与WT或 GmCDK8-OX 的总蛋白提取物孵育。B, C) 在本氏烟草 (N. benthamiana) 中共表达GmABI5-GFP和GmCDK8时的荧光强度和蛋白丰度分析。D-F) 双荧光素酶报告基因试验检测flg22处理下GmABI5对 GmCDK8 启动子的转录激活作用。G) 病原菌侵染诱导的依赖于GmABI5的 GmCDK8 转录激活。
3.6 GmABI5-GmCDK8模块抑制木质素生物合成途径
靶标高度重合:联合转录组学分析揭示,受GmABI5和GmCDK8调控的差异表达基因中有88%表现出一致的表达趋势,表明两者汇聚于相似的下游网络。
调控修饰通路:共同上调的基因主要富集于蛋白质复合物寡聚化、泛素-蛋白转移酶活性以及蛋白激酶活性等途径,暗示该模块广泛参与翻译后修饰。
抑木质素合成:共同下调的基因显著富集于次生细胞壁生物发生和苯丙素生物合成途径,特别是24个关键的木质素合成基因被协同抑制。
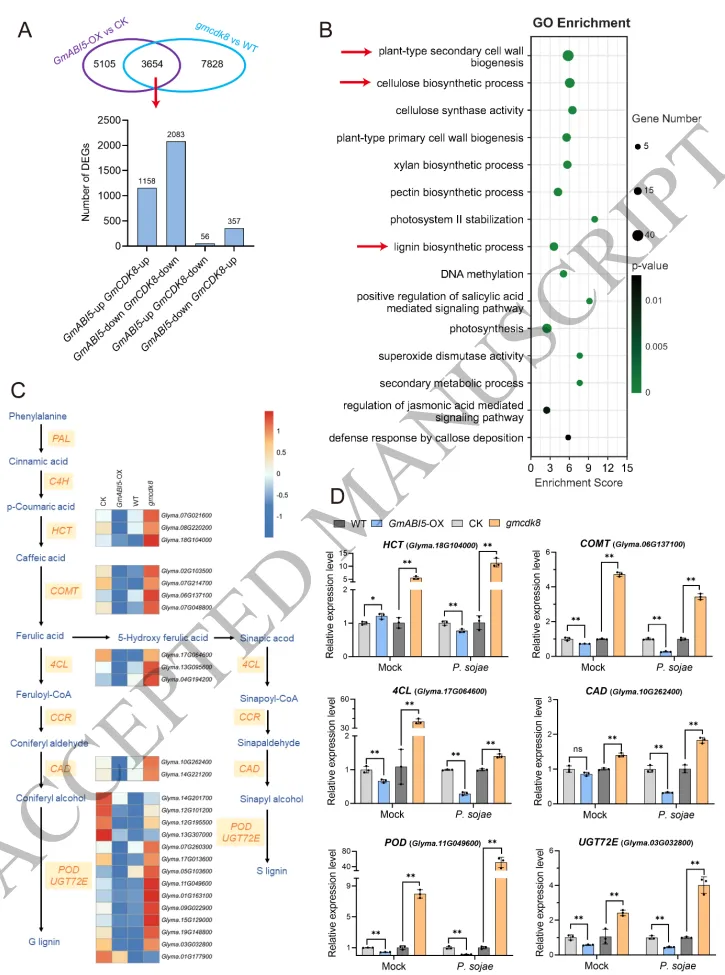
为了阐明GmABI5-GmCDK8模块在病原菌侵染期间激活的下游调控网络,联合转录组分析显示两者调控的差异表达基因中高达88%具有一致的表达趋势,证实它们协同作用于相似的靶标网络。功能富集分析表明,共同上调的基因多与泛素化和磷酸化等翻译后修饰过程相关;而共同下调的基因则高度集中在次生细胞壁形成和苯丙素生物合成途径。特别是包括 HCT、COMT、4CL、CAD 和 POD 等在内的24个木质素合成核心基因的表达受到了显著抑制,这一结果也得到了RT-qPCR的验证。这些数据充分表明,GmABI5-GmCDK8模块通过负向调控木质素生物合成途径,破坏了植物界面的物理防御,从而促进了大豆疫霉的侵染 (Fig. 6)。
图6. GmABI5-GmCDK8模块抑制木质素生物合成途径。 A) 维恩图显示受GmABI5和GmCDK8共同调控的差异表达基因 (DEGs) 的重叠情况及一致的表达趋势。B) 共同下调的DEGs的基因本体论 (GO) 富集分析,主要富集于次生细胞壁生物发生和木质素生物合成过程。C) 参与木质素生物合成途径的24个关键调控基因(包括 HCT、COMT、4CL、CAD、POD 和 UGT72E)的表达模式热图。D) RT-qPCR验证木质素生物合成相关核心基因的表达模式。