常规干燥过程中由含水率梯度及干缩异向性引起的干燥应力是导致木材干燥流变的主要原因,由毛细张力引起的皱缩是木材干燥流变的一种特殊现象,木材干燥流变应该涉及皱缩的研究。综述了木材干燥过程自由干缩、弹性、黏弹性蠕变及机械吸附蠕变应变的特征及微观响应机理,总结了4种应变的试验测量及模型预测方法,并对易皱缩木材的干燥流变特征进行了梳理。木材皱缩受干燥应力(拉/压)影响而出现回复/加剧,但其干燥流变规律和机制尚不明晰。因此,未来研究应采用先进的成像和数字相关(DIC)技术,深入探索干燥过程应力及各应变的产生、发展及回复的木材微观多尺度结构的响应机制。此外,应结合有限元和数值分析等方法研究细胞皱缩的干燥流变特性,从微观角度阐明毛细张力与干燥应力耦合作用下木材干燥过程中各阶段皱缩和收缩的流变发展机理。
木材是天然、绿色、环保材料,强重比高、弹性好、耐冲击且具有美观的纹理和色调。木材中的水分会影响其物理力学、加工性能及制品的稳定性。木材干燥是确保木材及制品品质的重要工序。木材干燥时,当含水率降至纤维饱和点(FSP)以下应产生自由干缩。但因木材较厚或干燥速率较快,实际干燥过程受到含水率梯度和干缩异向性产生的干燥应力共同作用导致非自由干缩,当干燥应力超过木材横向抗拉强度时,容易产生表裂、内裂等缺陷。
木材流变学研究木材在应力作用下变形随时间变化的特性,而木材干燥流变主要研究干燥过程中木材应力和应变的产生、发展及回复等变化特征与规律。传统木材干燥应变分为含水率应变和残余应变两类。含水率应变又称弹性应变,由含水率梯度应力引起且随含水率梯度的减小而降低,应力消失后可完全恢复。
当干燥应力超过木材比例极限或在比例极限范围内长时间作用时,应变在应力除去之后并未消失,为残余应变,存在于干燥结束后的木材中。
木材流变学理论引入木材干燥后,将干燥应力作用下的总应变分为自由干缩应变、瞬时弹性应变、黏弹性蠕变和机械吸附蠕变应变。弹性应变为瞬时发生,黏弹性蠕变随时间变化,自由干缩应变和机械吸附蠕变应变均受含水率变化的影响。在理想条件下,自由干缩过程没有含水率梯度应力,与含水率呈线性关系。但实际干燥过程由于含水率梯度难以消除,木材在应力作用下进行非自由干缩,作为弹塑性体的木材便会产生瞬时弹性应变、黏弹性蠕变和机械吸附蠕变。
某些树种的木材在干燥过程中除了产生常见的开裂、变形等缺陷,还极易产生不规则皱缩。木材皱缩与正常干缩不同,通常发生在FSP以上,因自由水迁移产生的毛细管张力所致。
目前,大多数研究将此类木材的变形称为全收缩,即干燥过程中皱缩和正常干缩之和,但对易皱缩木材干燥流变特性的研究很少。
研究木材干燥流变规律对实际生产中优化干燥工艺、提高干燥效率和干燥质量具有重要理论和实践意义。笔者综述了木材干燥流变特性的研究进展,对木材干燥应变实际测量及计算方法进行了系统梳理,并总结了易皱缩木材的干缩和流变特性,以期为木材干燥流变的研究提供参考。
1.1 干燥应变分类
1.1.1 自由干缩应变
自由干缩通常发生在FSP以下,为木材的固有特性。Zhan等发现自由干缩应变随温度的升高而减小,且在不同温度下从边材到髓心均呈现减小的趋势。微观上,木材收缩受到细胞壁组织、细胞形状、细胞类型、细胞数量、微纤丝角和孔隙率等影响。木材干燥过程中尺寸变化是由细胞壁中水分的损失引起的,水分子通过氢键结合到吸附位点(纤维素无定形区、半纤维素和木质素的羟基位点)。当细胞壁发生水的解吸时,细胞壁和整体细胞结构产生收缩,从而引起宏观尺度的收缩。Almeida等测量了挪威云杉(Picea asperata)早、晚材横向收缩,认为细胞层面的收缩是由细胞壁局部各向异性和形成层中细胞形状耦合造成的。Gao等在研究马尾松(Pinus massoniana)的收缩行为时发现弦向收缩受早、晚材管胞相互制约,但以晚材管胞收缩为主,而径向收缩因受射线组织约束,宏观收缩小于早、晚材收缩的均值。Perr等研究了挪威云杉滞后吸附膨胀/收缩行为,发现弦向和径向膨胀/收缩应变随孔隙率增加而增加。
在试验研究中,常用薄小试片在气干环境中测定自由干缩。Chen等制备了3 mm厚(纤维方向,L)赤桉(Eucalyptus camaldulensis)试片来测量自由干缩,因含水率梯度较小,产生的应力通常被忽略。自由干缩应变(εf)可由式(1)计算:
式中:α为木材自由干缩系数; M和MFSP分别为FSP以下和FSP时的木材含水率,M<MFSP。
1.1.2 受限干缩应变
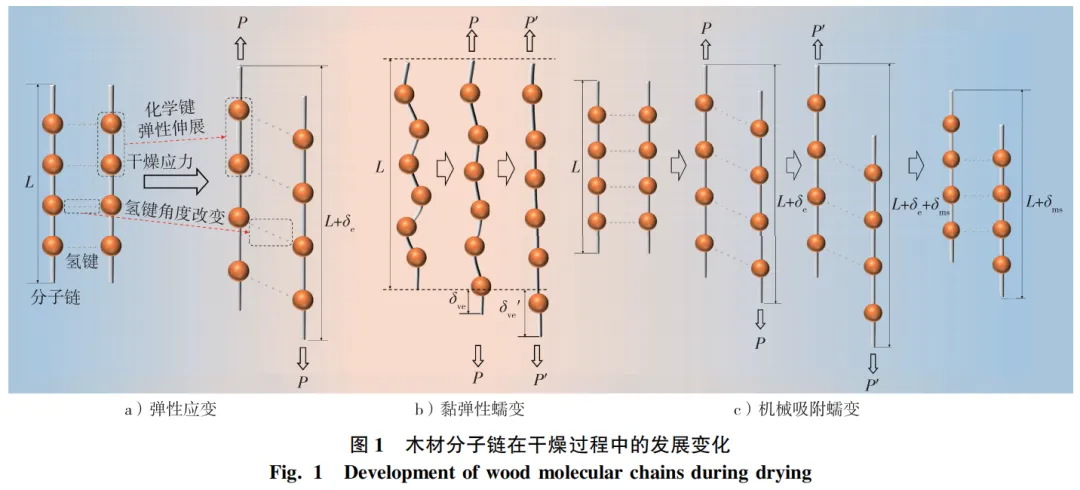
木材实际干燥过程通常经历的是受限干缩,即在应力作用下产生弹性应变、黏弹性蠕变应变和机械吸附蠕变应变。弹性应变反映木材内部的干燥应力,在受力瞬间产生,外力解除时立即复原,应力-应变关系满足胡克定律,微观上表现为一类和二类化学键弹性变形或键角变化(图1a)。
瞬时弹性应变(εe)可由式(2)计算:
式中:σ为木材所受应力,MPa; E为木材弹性模量,MPa。
蠕变是材料在应力作用下随时间变化的变形,包括黏弹性蠕变和机械吸附蠕变。黏弹性蠕变应变产生于没有水分变化的恒定干燥条件下,当载荷保持不变时,产生瞬时弹性响应后变形逐渐增加,而速率逐渐递减。去除载荷后,变形缓慢恢复至原来的形状。黏弹性蠕变应变可认为是分子链的卷曲,无外力时分子链自由能最低并保持较大的自由度。在外力作用下,分子链张紧并导致少量二类化学键断裂及重组(图1b)。在很多研究中,黏弹性蠕变应变因数值较小而被忽略不计或归属于机械吸附蠕变。黏弹性蠕变应变(εve)由式(3)计算:
式中:t为受载时间,min; a、q、n均为系数。
应力作用下因含水率变化导致木材变形率加大,这种现象称为机械吸附蠕变。与黏弹性蠕变不同,机械吸附蠕变发生于非恒定的干燥条件下,木材在应力下经历湿度或温度变化时产生。Chen等发现机械吸附蠕变的变化滞后于含水率变化,受瞬时干燥应力和干燥历史影响。机械吸附机理在于分子链间氢键的断裂和重组(图1c),在外力作用下分子链上水分子发生移动形成低能量结构,宏观表现为不可恢复的塑化变形。Haslach研究认为机械吸附行为是异质纤维中不均匀水分吸附所引起的含水率梯度。机械吸附蠕变(εms)由式(4)计算:
式中:m为机械吸附系数,MPa-1; ΔM为含水率变化量,%。
1.2 干燥应变测量方法
1.2.1 试验测算
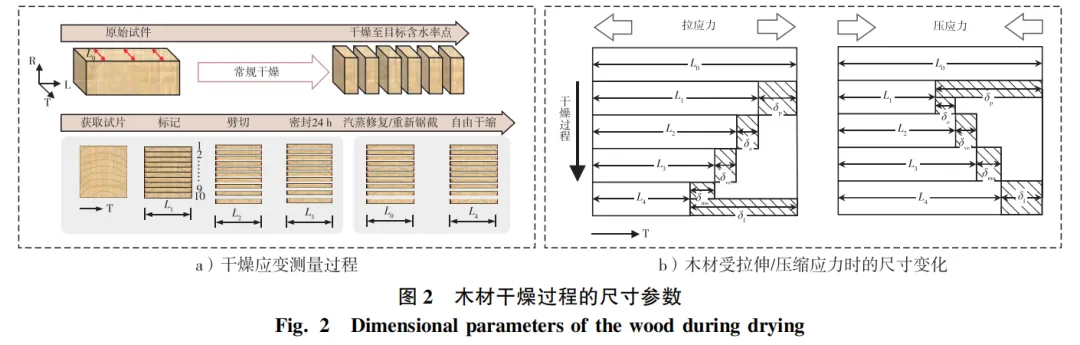
试验中一般采用分层切片法测量干燥应变,通过测量不同干燥状态下试片的长度L来确定弹性、黏弹性蠕变、机械吸附蠕变及自由干缩的应变值(图2、表1),具体测量方法如下:
1)干燥前,在应变检验板端部划线,测量弦向初始长度L0;
2)将应变检验板放入烘箱或恒温恒湿箱中进行干燥,干燥至目标含水率时取出试件,去掉端头并垂直于纤维方向锯截应变试片,在端面分层划线并测量各层长度L1。
3)用劈刀沿划线快速劈切各层,及时测量各层长度L2并称质量。
4)将劈切后试件用塑料薄膜密封(防止水分流失),在室温下使其自然恢复24 h,测量各层长度L3。
5)L4的测量有2种方法。第一,将所有试件用水浸泡24 h,常压饱和汽蒸8 h后在室温下自由干缩,其间定期测量各试件质量,当其质量与浸泡前质量相同时,测量各层长度L4(泡水-汽蒸)。第二,在同一检验板上应变试片相邻位置处锯截薄试片,分层划线并劈切后置于(25±3)℃、(70±5)%相对湿度(RH)下自由干缩,测量其到达目标含水率时的长度L4(直接切片)。研究发现,泡水-汽蒸并不能完全恢复试件前期干燥产生的塑化变形。而直接切片法极大削弱了干燥应力及皱缩的影响,直接切片法测量的自由干缩值更准确。
1.2.2 数值模拟
试验测量是获得具体数据最直接的方法,但仍存在人工误差等问题,目前很多研究通过建立模型来预测干燥应力和应变的发展。流变过程虽具复杂性,但其过程可通过弹簧和阻尼器的组合来描述,Maxwell和Kelvin-Voigt模型是描述黏弹性行为的2种典型力学模型。标准弹簧与阻尼器串联组合形成Maxwell模型,而弹簧与阻尼器并联组合成Kelvin-Voigt模型。Maxwell和Kelvin-Voigt模型串联得到Burger模型,虽也用于描述黏弹性,但不能完全解释木材中产生的干燥应力。
传统的黏弹性流变应力模型不足以模拟不同含水率下的干燥应力,在干燥应力模型中必须考虑机械吸附效应。Tu等在木材干燥流变的基础上建立了干燥应力模型,可直接通过测量木材各层实际收缩率和含水率来计算应力。Moutee等的流变模型可预测干燥过程中应力反转、残余应力和厚度方向的最大应力。Pérez-Pena等使用有限元法模拟了桉木干燥过程中由含水率变化和机械吸附效应所产生的干燥应力。Hanhijärvi等在干燥应力一维模型的基础上,引入塑性单元扩展为二维模型描述部分不可恢复的机械吸附蠕变,研究木材干燥的黏弹性-机械吸附塑性力学行为。Zhu等构建了热质传递耦合数值模型模拟了干燥过程中湿热耦合对木材收缩变形的影响及应力反转现象。Florisson等构建了三维数值模型,通过有限元法模拟了含水率影响下木材的变形和应力。
人工神经网络(ANN)作为近年来较热门的人工智能算法在木材干燥领域也有应用。付宗营将ANN用于干燥应变的模拟预测,建立了预测弹性应变和机械吸附蠕变的三层前馈型网络模型,证实94%的试验数据有效。Fu等还提出了用于预测白桦(Betula platyphylla)圆盘在干燥过程中受温度、含水率、相对湿度和髓心距影响的弹性应变的ANN模型; 之后又预测了经预蒸后白桦圆盘的机械吸附蠕变,R2均大于0.95,相关性很高。由于采用反向传播神经网络(BPNN)无法预测下一时刻干燥应力的发展,Gao等先利用GM(1,1)灰色预测模型对未来相邻时刻含水率进行预测,在此基础上构建了BPNN来预测表、心层弹性应变的发展趋势。
木材干燥流变特性受木材结构、干燥时间、含水率、水分历史、温度、湿度、干燥应力等内外因素影响。木材从宏观至微观的多级组织构造作为内因始终影响其干燥特性,而可人为干预的干燥时间、温度、湿度和应力等外因也对木材干燥特性及干燥质量有重要影响。应力是导致干燥应变的直接原因,当应力水平超过弹性极限时可观察到黏弹性蠕变和机械吸附蠕变。
2.1木材组织结构的影响
由于生长环境及树种不同,木材在组织构造上会存在较大差异,即使是同一树种木材,若取材位置不同,基于细胞水平的细胞形状、长度、直径、细胞壁组织、纤维素、半纤维素及木质素等多级构造也不同,材性均会有较大差异。刁海林等指出,马尾松的抗瞬间弹性变形、抗长期黏性蠕变及抗延迟弹性蠕变能力均比尾赤桉强。尹业桥等研究表明,早材管孔带的存在对于环孔材栎木(Quercus spp.)蠕变应变具有较大影响。Navi等指出,压缩木材的纤维比普通木材纤维黏弹性更强,这主要由压缩木材次生细胞壁(S2层)和成材早期木材微纤丝角不同所决定。毕俊杰等研究发现软木细胞在横切面和径切面呈现砖块状排列,在弦切面为蜂窝状排列,这种独特的细胞结构和生长方式导致软木动态黏弹性存在各向异性。初生栓皮栎软木的刚性与黏性均强于再生栓皮栎软木,抵抗变形的能力更强。Zhang等研究了去木质素处理对木材机械吸附蠕变的影响,发现去木素化对木材径向的影响更大。王聪等研究了不同纹理方向栎木微小无疵试样板材蠕变特性,发现栎木板材横向比纵向更易发生黏性形变。
2.2温度和含水率的协效影响
木材作为一种非均质细胞结构生物聚合物黏弹性材料,其力学性能取决于温度和含水率。干燥过程中动态变化的温度和含水率对木材的流变具有协同作用,但温度变化对蠕变的影响可能被含水率变化引起的机械吸附蠕变所覆盖。Lagana等通过Burger模型研究了挪威云杉的流变参数,发现4个流变参数均随温度从120 ℃升高至180 ℃而降低,温度升高会导致弹性变形、变形速率及蠕变增加。这主要由高温下木质素的塑化以及半纤维素的解聚所致。但Zhan等的研究中发现,木材表面的黏弹性和机械吸附蠕变随干燥温度从40 ℃升高到80 ℃而下降。
含水率影响木材的力学强度、弹性模量和蠕变等性能。当木材含水率低于FSP时,弹性模量随含水率的增加而降低。弹性模量的降低使木材强度降低,从而增加弹性变形和正常蠕变。Armstrong等研究了含水率变化对木材流变特性的影响,结果显示含水率的变化会影响变形的速度和幅度,形变大小主要取决于含水率变化的程度而非速率。虽然黏弹性蠕变主要受时间影响,但黏弹性特性依赖于含水率。在干燥早期高含水率阶段,黏弹性蠕变会增大并加快变形速度。但随干燥的持续,黏弹性蠕变的发展因含水率降低可忽略不计。Fu等研究发现弹性应变和黏弹性蠕变均随着含水率的降低而减小,并且在相同含水率水平下弦向应变高于径向。受含水率及累积蠕变的影响,机械吸附蠕变的发展与收缩量并不成正比,当收缩增加时,蠕变发展会略有减少。Hanhijärvi等在恒定试验条件下观察到蠕变对温度和含水率非常敏感,高含水率木材在高温下的蠕变比低含水率木材在低温下的蠕变更快。
2.3干燥应力的影响
剧烈干燥条件下产生的拉/压应力是导致机械吸附蠕变的根源。拉/压应力贯穿于整个干燥过程,并在不同时期可相互转换。干燥前期,表层由于干燥较快而产生收缩应变,心层因含水率较高未产生收缩而限制了表层的收缩,表层受到来自心层的拉应力,心层则受到相反的压应力。此时,木材表层因机械吸附效应而产生了永久变形,并限制了木材自由干缩变形。在干燥后期,应力模式发生逆转,表面处于压缩状态,而心层处于拉伸状态。Fu等指出由于收缩各向异性,弦向收缩应变受径向收缩限制。因此,弦向产生拉应力,径向产生压应力。弦、径向收缩率相同时则不存在内应力,一旦径向收缩率超过弦向就会发生应力反转。Rice等研究了红橡木(Quercus rubra)机械吸附蠕变发展过程,发现干燥试件表面应力反转时机械吸附蠕变最大,当拉应力反转为压应力时,则会减小机械吸附蠕变的幅度。
战剑锋等研究了白桦木干燥过程的横纹流变特性,发现机械吸附蠕变沿厚度方向随湿热处理过程的延续有所增长,认为机械吸附效应是木材干燥内应力释放的主要原因。Zhan等提出了“机械吸附蠕变梯度”概念描述常规干燥过程中干燥应力和应变的发展,定义了干燥应力和机械吸附蠕变变量之间的定量公式,可通过测量含水率、收缩率和机械吸附蠕变定量确定窑干木材的应力状态。Ormarsson等研究发现,当存在机械吸附效应时产生的应力低于仅有弹性应变时产生的应力,机械吸附效应对应力的影响较弹性应变更大。Langrish研究发现,由于机械吸附蠕变在干燥初始阶段和整个干燥期间释放了应力,因此产生了较低的瞬时弹性应变。Kang等通过数学模型预测了树盘中的干燥应力和裂纹形成,发现干燥应力在机械吸附效应的影响下随含水率的降低呈非线性变化,树盘的正交各向异性越高干燥应力越大。Pang研究发现通过频繁控制气流反转可以降低干燥过程中的最大应力及残余应力。空气湿度和温度循环改变了木材含水率,从而导致机械吸附蠕变增加,内应力得到释放,降低了木材开裂变形的风险。对于存在较高内应力的木质产品,可利用木材流变特性和湿度变化的方法,将残余应力降低到可接受的水平。
3.1木材皱缩的机制
水分在木材内部以细胞壁中的吸着水、管腔及孔隙中的自由水或水蒸气3种形式存在。木材干燥过程中,自由水的束缚力较小而蒸发较快,在纹孔膜微孔处产生较大的毛细张力,从而导致易皱缩木材在FSP以上时发生异常收缩。木材产生皱缩需具备3个基本条件:①管腔饱水且具良好气密性; ②在弯液面处产生毛细管张力; ③垂直于细胞壁的平均抗压强度小于毛细管张力诱导的应力。此外,当木材表层含水率低于FSP时产生的含水率梯度应力会作用于细胞壁,影响木材的皱缩。因此,木材干燥皱缩需从微观结构出发,研究细胞壁、木材组分等的干缩过程及与水分的作用机理。
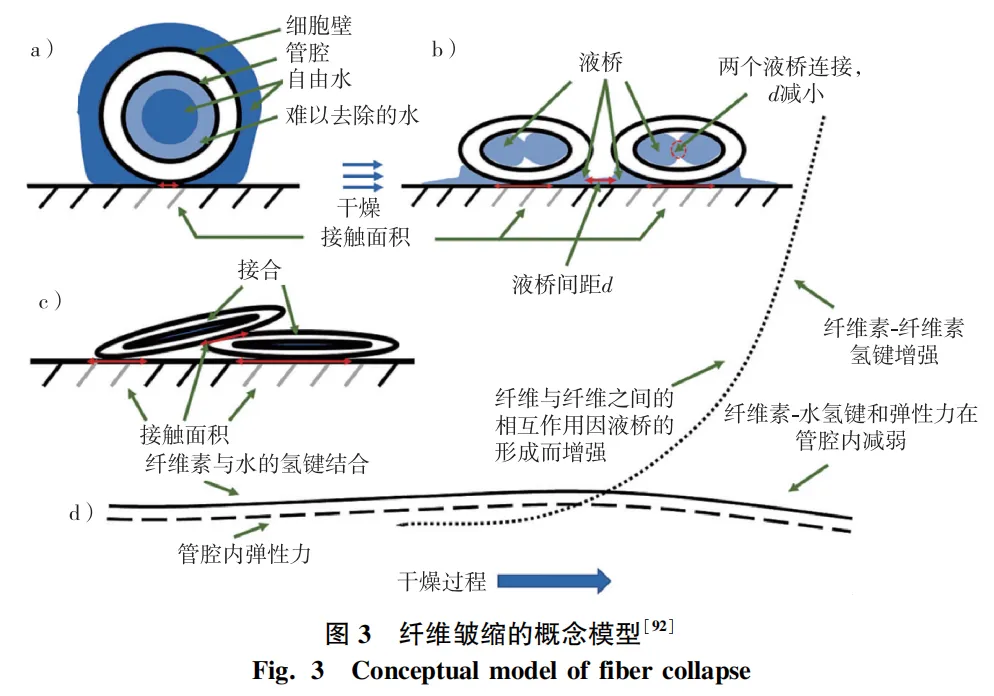
木材主要化学成分是构成木材细胞壁和胞间层的物质,主要由纤维素、半纤维素和木质素组成,占木材总量的90%以上。纤维素作为细胞壁骨架物质,与干燥过程中的收缩、皱缩密切相关。Salem等详细阐释了水分与纤维皱缩的关系,认为细胞中还存在另一种形式的截留水,干燥时被限制在纤维网格几何形状中(图3)。这种水介于吸着水和自由水之间,将吸着水和截留水称为“难以去除的水”(HR水)。由于HR水在干燥中更难去除,会在管腔内部和周围纤维细胞之间形成液桥(图3b)。随着HR水的去除,毛细张力使纤维细胞之间的距离更近,纤维素-水之间的氢键断裂,并形成纤维素-纤维素氢键(图3d)。纤维之间的接触面积随干燥时间延长而增大,管腔内壁压力降低,从而降低了纤维细胞管腔的弹性,最终导致不可逆的细胞壁变形和孔隙闭合,即“纤维细胞皱缩”。Akbari等通过建立管腔和细胞壁微孔的弹性毛细管模型研究发现,当弯液面长度超过微孔直径或管腔宽度时,所产生的毛细张力并不足以克服细胞壁拉伸阻力,因此不太可能导致皱缩。
3.2易皱缩木材的干燥流变
3.2.1 自由干缩特性
易皱缩木材自由干缩的测量可通过控制试片厚度来实现。陈太安用5 mm(L)厚赤桉试片作为无皱缩对照组在(20±2)℃,(65±3)%RH条件下气干测试自由干缩。Zhang等同样利用5 mm(L)厚尾巨桉(Eucalyptus urophylla × E. grandis)试片在(25±3)℃、(70±5)%RH条件下进行测试,但发现在FSP以上出现了明显的皱缩。杨琳等制备了1,2,3 mm(L)厚尾巨桉试片,在25 ℃、98%RH的环境中研究其干缩特性,发现即使是1 mm厚试片在高含水率范围内也产生了皱缩,无法实现自由干缩。木材纤维的平均长度为0.6~0.8 mm,若沿纤维方向制备试片的厚度小于0.6 mm,则可切断大部分木材纤维,使纤维细胞失去密闭性,毛细张力将不会形成,可从根本上消除木材皱缩。Redman等发现,由于出现皱缩,1 mm厚试片也无法表征柠檬桉(Corymbia citriodora)的径向和弦向收缩(27 ℃,88%RH); 他们尝试利用0.5 mm厚试片表征无皱缩的收缩,结果表明,在FSP以上时0.5 mm厚试片的皱缩最小,但1和2 mm厚试片则出现了明显皱缩,因此,利用0.5 mm厚试片的含水率-收缩率曲线拟合了FSP。易皱缩木材试片越厚则越容易产生皱缩,从而影响FSP的拟合精度。Dawson等使用超临界CO2对斜叶桉(Eucalyptus obliqua)进行预处理,之后制备了0.8和5.0 mm厚试片分别研究无皱缩和纯皱缩收缩,发现超临界CO2预处理木材的皱缩在25 ℃,65%RH的环境中极大减少。但Almeida等发现0.5~2.0 mm厚山毛榉(Fagus longipetiolata)试片干燥至20%~40%含水率时均产生了相同趋势的异常收缩,他们认为与射线细胞皱缩相关,射线细胞平均长度为30~80 μm,因此不会被切断。
3.2.2皱缩对干燥应力的影响
皱缩是干燥早期自由水迁移引起的毛细张力所致,随着木材表层含水率降至FSP以下,吸着水迁移会产生干燥应力,因此,易皱缩木材的流变是在毛细张力和干燥应力的耦合作用下产生的。已有大量研究指出,由干燥产生的拉/压应力直接影响干燥应变的大小,但是否会对干燥前期已产生的皱缩造成影响尚不清楚。Blakemore指出,如果皱缩发生在FSP以上,则可能在一定程度上降低含水率梯度对正常干缩的影响,且表面发生的皱缩可能会加剧后续的皱缩,并深入板材内部,即皱缩的发生极大增加了干燥应力和开裂的可能性。Pérez-Peña等研究了桉木在干燥过程中产生的机械变形和总挠度,认为可以利用机械吸附行为来减轻皱缩和干燥应力。王喜明等对比了3种桉木的干燥皱缩特性,认为在产生干燥皱缩后,干燥应力会加剧皱缩细胞的皱缩程度,毛细张力和干燥应力均为皱缩的产生条件,且其贡献率分别为75%和25%。陈振兴研究发现,拉应力可降低木材皱缩量,0.4 MPa的压应力可完全抑制皱缩恢复。




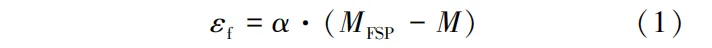
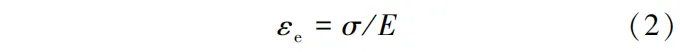
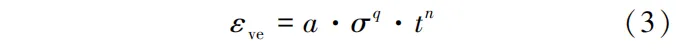
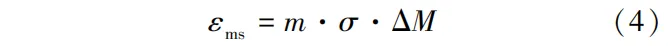








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
