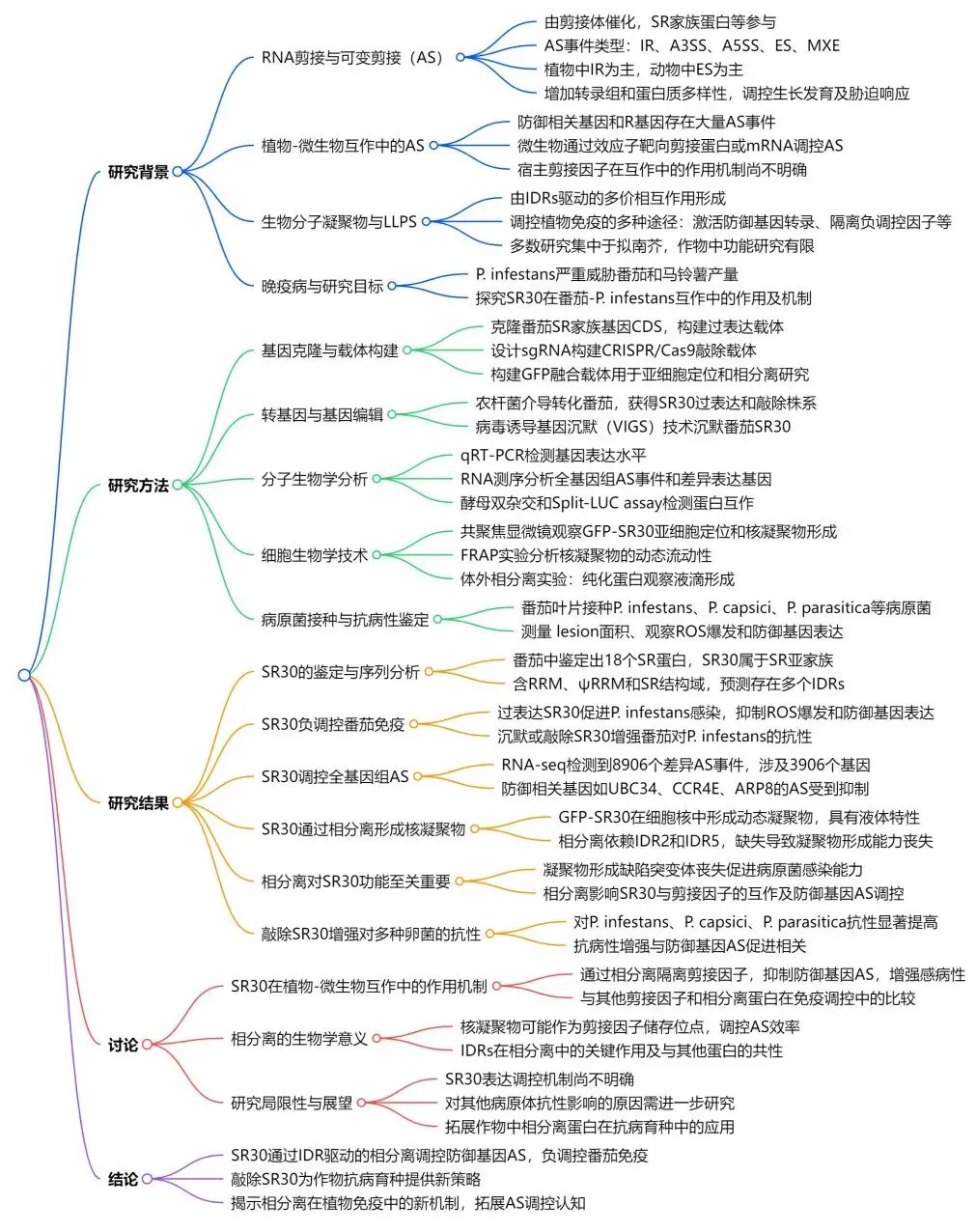
为了进一步研究SR30在植物免疫中的功能,构建稳定过表达SR30的转基因番茄(SR30-OE)。检测疫霉菌感染后ROS爆发和防御基因(如LRR22、GRAS2、WRKY28等)的表达。发现与WT植株相比SR30-OE植株ROS水平降低,防御基因表达下调,病斑面积增大。随后在番茄植物中进行SR30的瞬时沉默,接种测定显示SR30的沉默显著增强番茄对致病疫霉的抗性(图1d-k)。这些结果说明SR30负调控番茄免疫,抑制防御反应。
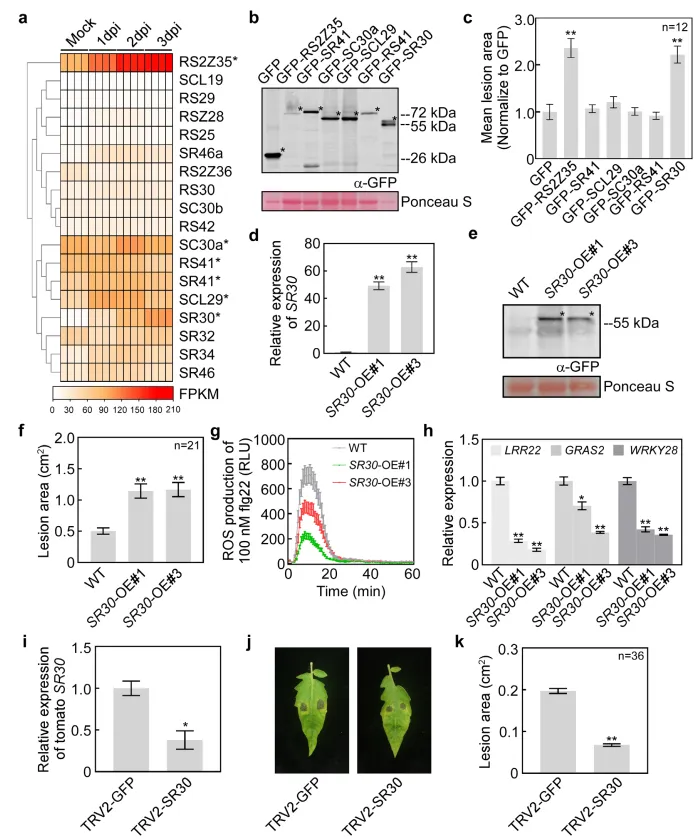
SR家族蛋白对于前体mRNA剪接的调节是重要的。为了揭示SR30如何通过剪接调控影响免疫,研究人员对过表达SR30-OE和野生型WT番茄进行RNA测序(RNA-seq),分析差异剪接事件(DASE)。结果表明SR30过表达导致8906个差异剪接事件,涉及内含子保留(IR)和外显子跳跃(ES)等类型,通过比对发现差异剪接基因(DASGs)与差异表达基因(DEGs)重叠较少(图2),表明SR30主要影响剪接而非转录。这些结果表明SR30通过调控全基因组mRNA剪接影响植物免疫。
图2. SR30调控番茄防御相关基因的可变剪接(AS)
a)使用Illumina平台进行转录组测序的示意图。采集SR30过表达(SR30-OE)和野生型(WT)番茄(Micro-Tom)叶片样本,提取RNA并合成双链cDNA文库。所有RNA-seq数据均进行差异基因表达和差异可变剪接分析。b)SR30-OE与WT比较中鉴定出的差异可变剪接事件(DASEs)数量及对应基因数。c)SR30-OE与WT比较中鉴定出的差异表达基因(DEGs)数量,“Up”和“Down”分别表示上调和下调的DEGs。d)维恩图显示SR30-OE与WT比较中差异可变剪接基因(DASGs)与DEGs的数量重叠关系。e-g)三个番茄防御相关基因的内含子剪接型和内含子保留型异构体剪接比率验证。左侧第一栏展示了三个选定基因的RNA-seq数据波动图:泛素结合酶34(UBC34,e)、碳代谢物阻遏蛋白4E(CCR4E,f)和肌动蛋白相关蛋白8(ARP8,g),并附两种不同转录本异构体的基因模型示意图。星号表示提前终止密码子(PTC)。左侧第二栏显示使用异构体特异性引物通过qRT-PCR检测三个候选基因两种异构体的相对转录水平。第三栏显示三个防御相关基因的剪接比率(内含子剪接型转录水平/内含子保留型转录水平),以番茄泛素基因UBI作为内参,相对转录水平以WT的内含子保留型异构体为基准进行标准化。数据表示为平均值±标准误(SE,n=3),P值通过Student's t检验分析(**,P<0.01)。最后一栏显示三种防御相关基因不同异构体在植物抗致病疫霉菌(P. infestans)免疫中的功能分析,以GFP作为对照,病斑面积在接种后5或6天(dpi)测量并标准化至GFP对照。数据表示为平均值±SE(n=20),P值通过Student's t检验分析(**,P<0.01)。
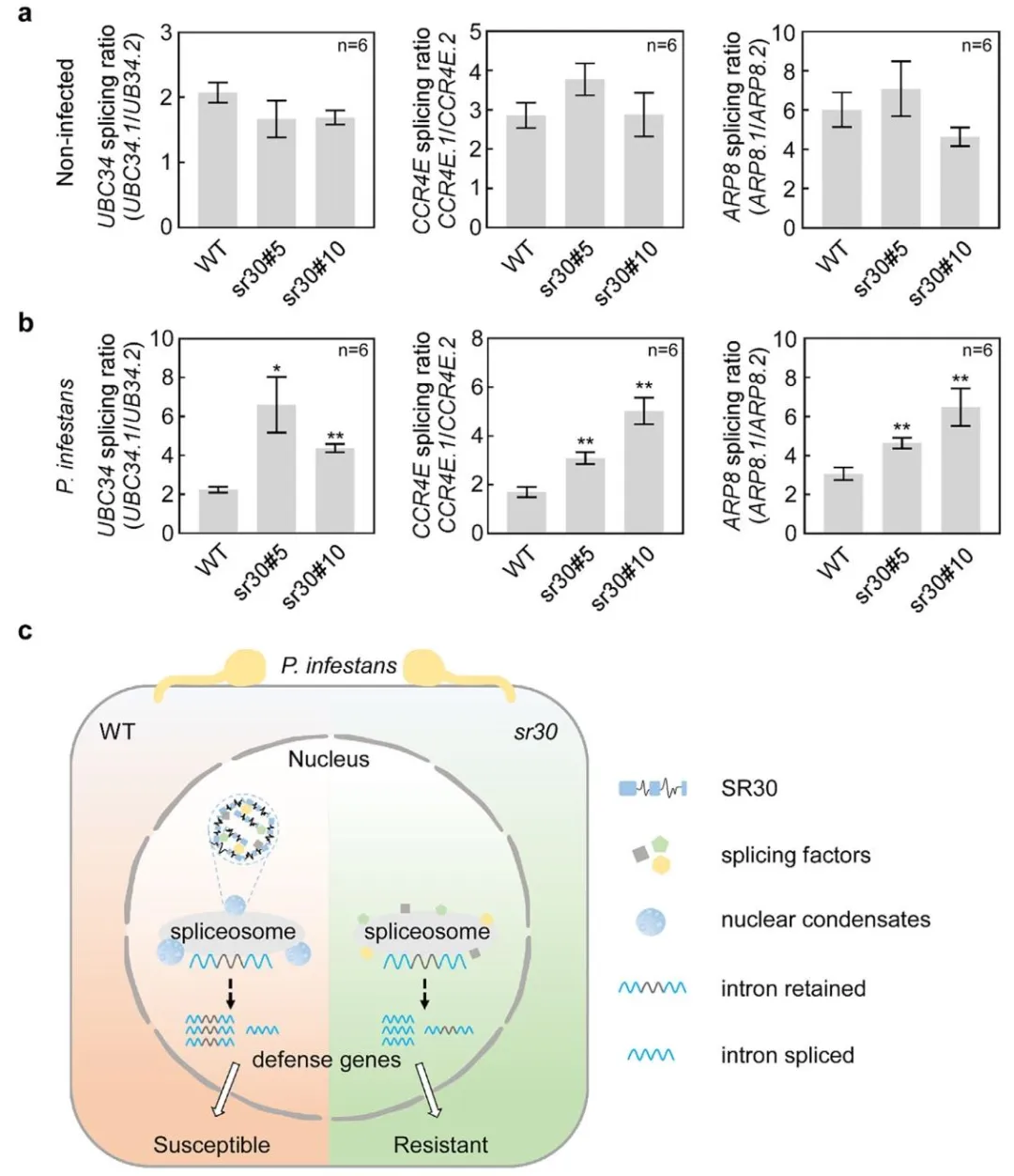
为了研究SR30如何通过剪接调节植物免疫,选择候选基因(如UBC34、CCR4E、ARP8),分析其剪接异构体的功能,同时在烟草中过表达不同剪接异构体,观察其对疫霉菌抗性的影响。结果发现功能性剪接体(如UBC34.1)抑制病原体生长,而内含子保留的剪接体(如UBC34.2)无功能。此外,与WT植株相比SR30-OE植株能降低功能性剪接体的比例(图2)。这些结果说明SR30通过抑制防御基因的功能性剪接,削弱植物抗病性。
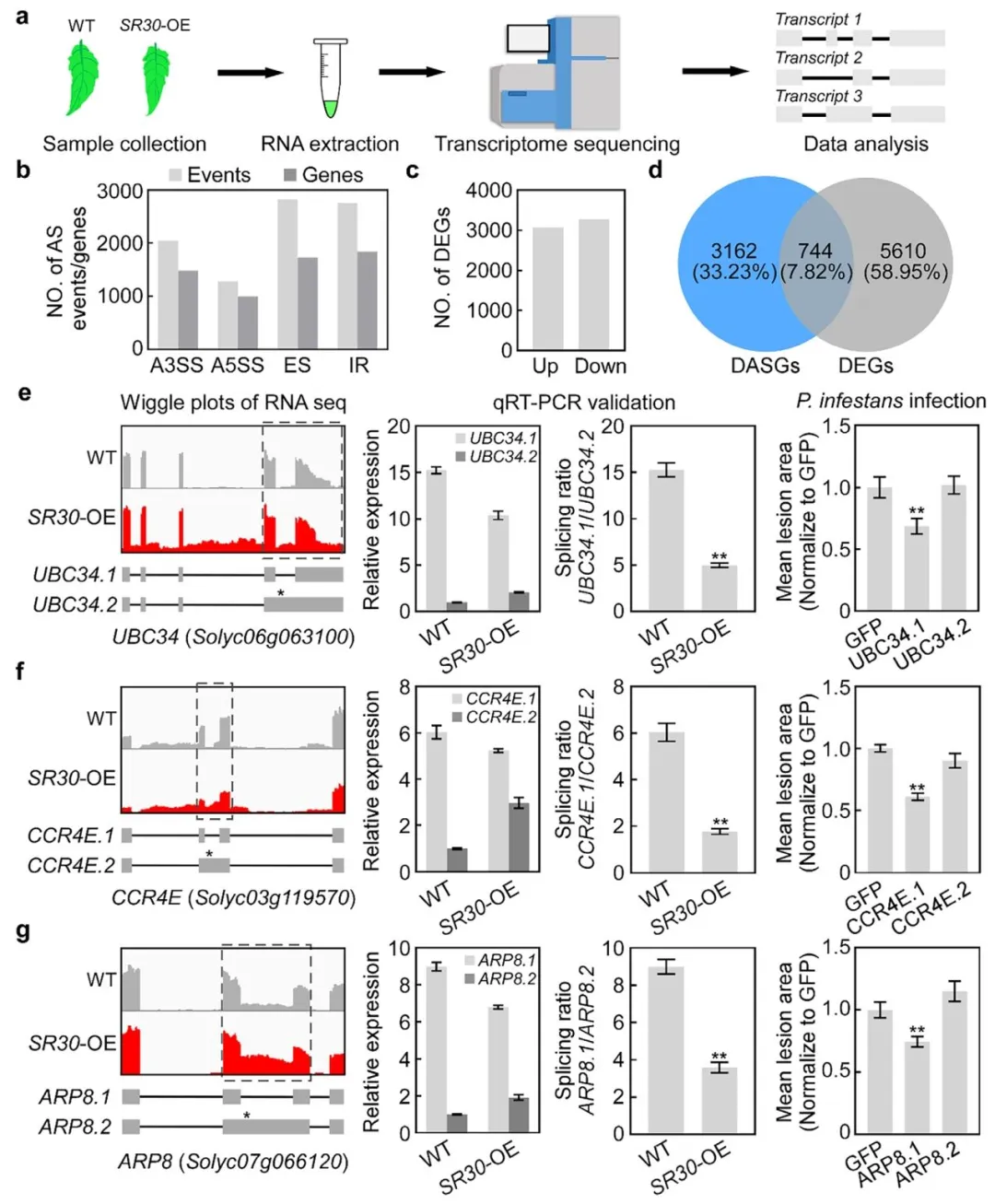
为了进一步探究SR30调控剪接的分子机制,通过荧光标记SR30(GFP-SR30),观察其在细胞核内的定位。结果显示,GFP-SR30形成各种大小的离散核凝聚物,而GFP在整个细胞核中不形成任何凝聚物;在SR30-OE番茄叶核中也观察到核浓缩物的亚细胞定位。此外,发现GFP-SR30能与拟南芥SR45共定位,AtSR45已被证明在体外剪接测定中作为剪接因子起作用,因此,推断SR30可能位于储存剪接因子的核斑点中。为了确定SR30蛋白的分子动力学,研究人员进行了荧光漂白恢复(FRAP)和体外液滴实验验证凝聚体的流动性。结果显示,GFP-SR30的核冷凝物在光漂白后一分钟内迅速恢复。此外,SR30的两个核凝聚体能够融合成一个凝聚体(图3)。这些结果表明SR30通过液-液相分离形成动态核凝聚体,可能作为剪接调控的枢纽。
图3. SR30通过液-液相分离(LLPS)形成核凝聚物
a)GFP和GFP-SR30的亚细胞定位。GFP和GFP-SR30在表达核标记物(H2B-RFP)的转基因本氏烟草(N. benthamiana)叶片中瞬时表达48小时后,通过共聚焦显微镜(20×和63×物镜)成像。比例尺为10微米。b)GFP-SR30在SR30过表达(SR30-OE)转基因番茄叶片中的亚细胞定位。比例尺为5微米。c)GFP-SR30在本氏烟草叶片表皮细胞中的荧光恢复后光漂白(FRAP)分析。白色箭头指示光漂白的核凝聚物。比例尺为10微米。d)光漂白后GFP-SR30的相对荧光恢复曲线。数据通过基于网络的工具easyFRAP进行标准化。数据表示为平均值±标准差(SD,n=10)。e)两个独立的GFP-SR30核凝聚物的融合过程。箭头指示正在融合的两个核凝聚物。比例尺为10微米。f)体外纯化蛋白GFP-SR30的液滴形成能力分析。上图为His-MBP-GFP和His-MBP-GFP-SR30重组蛋白的示意图,TEV蛋白酶用于去除His-MBP标签。下图为1 μM GFP-SR30在150 mM NaCl存在下的体外液滴形成图像,以GFP蛋白作为对照。比例尺为10微米。
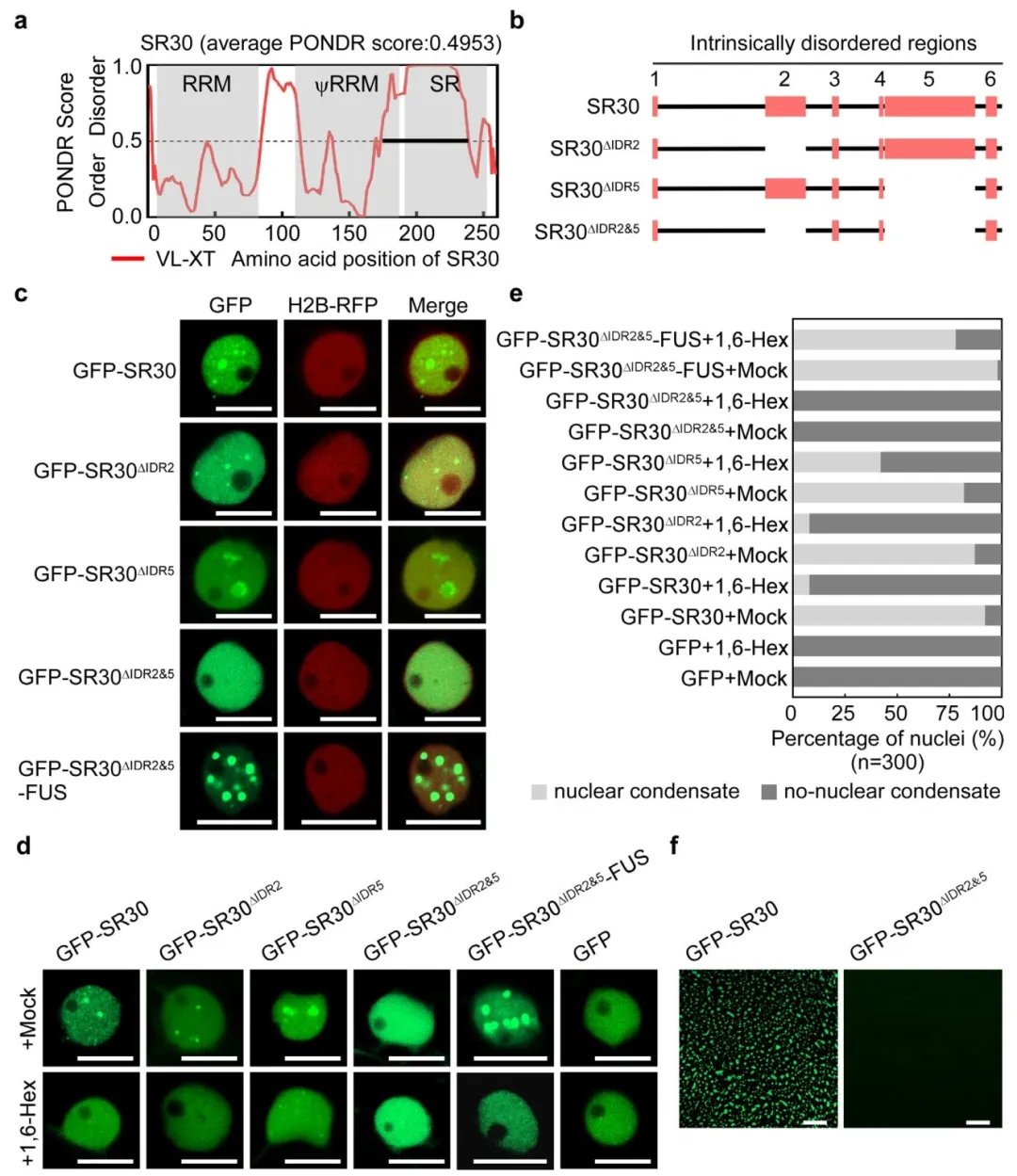
内在无序区(IDR)介导多价相互作用,是相分离的主要驱动力。为了确定驱动SR30相分离的关键结构域,研究人员预测SR30的IDR(IDR2和IDR5),构建IDR缺失突变体(GFP-SR30Δ IDR 2、GFP-SR30Δ IDR 5、GFP-SR30Δ IDR 2&5)。与GFP-SR30相比,GFP-SR30Δ IDR 2的核凝聚物显示出显著降低,而GFP-SR30Δ IDR 5显示出增加,只有GFP-SR30Δ IDR 2&5突变体不能形成核凝聚物。这说明IDR2和IDR5都是SR30在体内形成核冷凝物所必需的。随后在SR30Δ IDR 2&5-FUS的C端引入一个RNA结合蛋白FUSED IN SARCOMA(FUS)的低复杂性结构域(LCD)来挽救SR30Δ IDR 2&5的相分离,结果GFP-SR30Δ IDR 2&5-FUS重新形成核凝聚物(图4)。这些结果表明SR30的相分离依赖IDR2和IDR5,二者通过多价相互作用驱动凝聚体形成。
图4. SR30的液-液相分离(LLPS)由内在无序区域(IDRs)驱动。
a)使用PONDR预测SR30的内在无序区域(IDRs)。三个灰色框分别代表SR30的RNA识别基序(RRM)、假RNA识别基序(ψRRM)和丝氨酸/精氨酸富集(SR)结构域区域。b)三种设计的SR30 IDR缺失突变体的示意图。c)SR30的IDR缺失变体和互补突变体SR30^{IDR2&5}-FUS的亚细胞定位。所有突变体在表达核标记H2B-RFP的转基因本氏烟草叶片中表达48小时后,使用共聚焦显微镜成像。比例尺表示10微米。d)SR30的IDR缺失突变体和SR30^{IDR2&5}-FUS在1,6-己二醇(1,6-Hex)处理下的亚细胞定位。所有蛋白在本氏烟草中表达36-48小时。对于模拟处理或1,6-Hex处理,在观察前用 Milli-Q水或5% 1,6-Hex浸润叶片组织。比例尺表示10微米。e)与模拟对照相比,经1,6-Hex处理的IDR缺失突变体和互补突变体SR30^{IDR2&5}-FUS中含有核凝聚物的细胞核百分比。每种处理共计算300个细胞核。f)1μM GFP-SR30和GFP-SR30^{IDR2&5}在150 mM NaCl存在下的体外液滴形成。比例尺表示10微米。
为了探讨SR30的疾病易感性与其相分离能力之间的关系,研究人员在烟草中表达SR30突变体(GFP-SR30Δ IDR 2&5,GFP-SR30Δ IDR 2&5-FUS),发现与GFP相比,GFP-SR30Δ IDR 2&5丧失促进病害的能力,而GFP-SR30Δ IDR 2&5-FUS能恢复其功能。这一结果表明,由IDR介导的相分离是SR30的疾病易感性功能所必需的。为了进一步研究SR30如何抑制前体mRNA剪接,使用Split-LUC试验检测了SR30、SR30Δ IDR 2&5、SR30Δ IDR 2&5-FUS和8种剪接因子(SR30、SR41、SCL19、SCL29、RS29、RS30、SR46、U1-70K)之间的相互作用,结果发现SR30通过相分离与其中5种剪接因子(如SCL19、SR46)互作,SR30Δ IDR 2&5的相互作用会减弱,而GFP-SR30Δ IDR 2&5-FUS部分恢复了相互作用能力(图5)。这些结果说明SR30的相分离是其抑制剪接和增强病害易感性的必要条件。
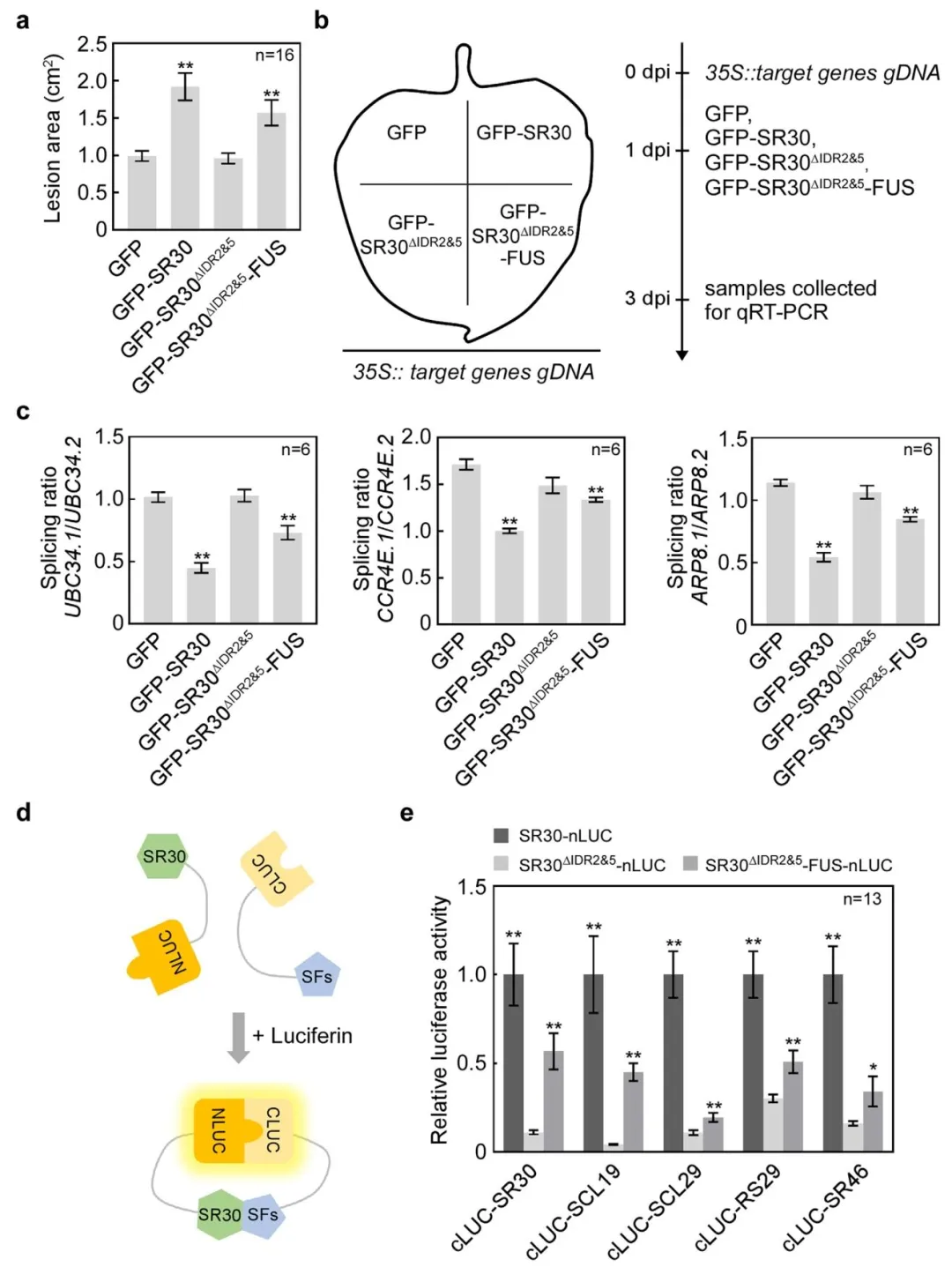
图5. SR30通过液-液相分离(LLPS)调控防御基因的可变剪接(AS)以发挥感病性功能。a)表达GFP、GFP-SR30、GFP-SR30ΔIDR2&5和GFP-SR30ΔIDR2&5-FUS的本氏烟草叶片在接种后5天(dpi)的病斑面积(cm²)。数据表示为平均值±标准误(SE,n=16),P值通过Student's t检验分析(**,P<0.01)。b)验证SR30相分离突变体对靶基因AS影响的流程示意图。步骤1:将携带35S::靶基因gDNA(35S::SlUBC34/SlCCR4E/SlARP8 gDNA)的农杆菌菌株浸润本氏烟草全叶;步骤2:在浸润后1天(dpi),于同一株表达35S::靶基因gDNA的本氏烟草叶片的不同区域分别表达GFP、GFP-SR30、GFP-SR30^{Delta IDR2&5}和GFP-SR30ΔIDR2&5-FUS;步骤3:分别收集表达不同蛋白的本氏烟草叶片,提取所有处理样品的总RNA并进行qRT-PCR检测。c)通过qRT-PCR分别检测GFP、GFP-SR30、GFP-SR30^{Delta IDR2&5}和GFP-SR30ΔIDR2&5-FUS处理下UBC34、CCR4E和ARP8两种不同异构体的剪接比率(内含子剪接型/内含子保留型)。以本氏烟草肌动蛋白基因(NbActin)作为内参,每个靶基因的不同处理组中两种异构体的相对转录水平均以内含子剪接型的转录水平为基准进行标准化。数据表示为平均值±SE(n=6),P值通过Student's t检验分析(**,P<0.01)。该实验重复两次,结果相似。d)分裂荧光素酶互补实验(split-LUC assays)检测SR30与其他剪接因子(SFs)相互作用的示意图。将SR30、SR30^{Delta IDR2&5}和SR30^{Delta IDR2&5}-FUS插入pICH86988-nLUC载体,其他剪接因子插入pCAMBIA1300-cLUC载体。如果两种待测蛋白相互作用,荧光素酶在荧光素存在下会发出荧光。e)通过split-LUC实验分别检测SR30、SR30^{Delta IDR2\&5}和SR30^{Delta IDR2&5}-FUS与其他番茄剪接因子的相互作用相对荧光素酶活性(LUC)。共表达目的蛋白48小时后,利用叶圆片测量发光强度。SR30^{Delta IDR2&5}和GFP-SR30ΔIDR2&5-FUS与各剪接因子的相对LUC活性均以SR30与相同剪接因子的相互作用活性为基准进行标准化。数据表示为平均值±SE(n=13),P值通过Student's t检验分析(*,P<0.05;**,P<0.01)。
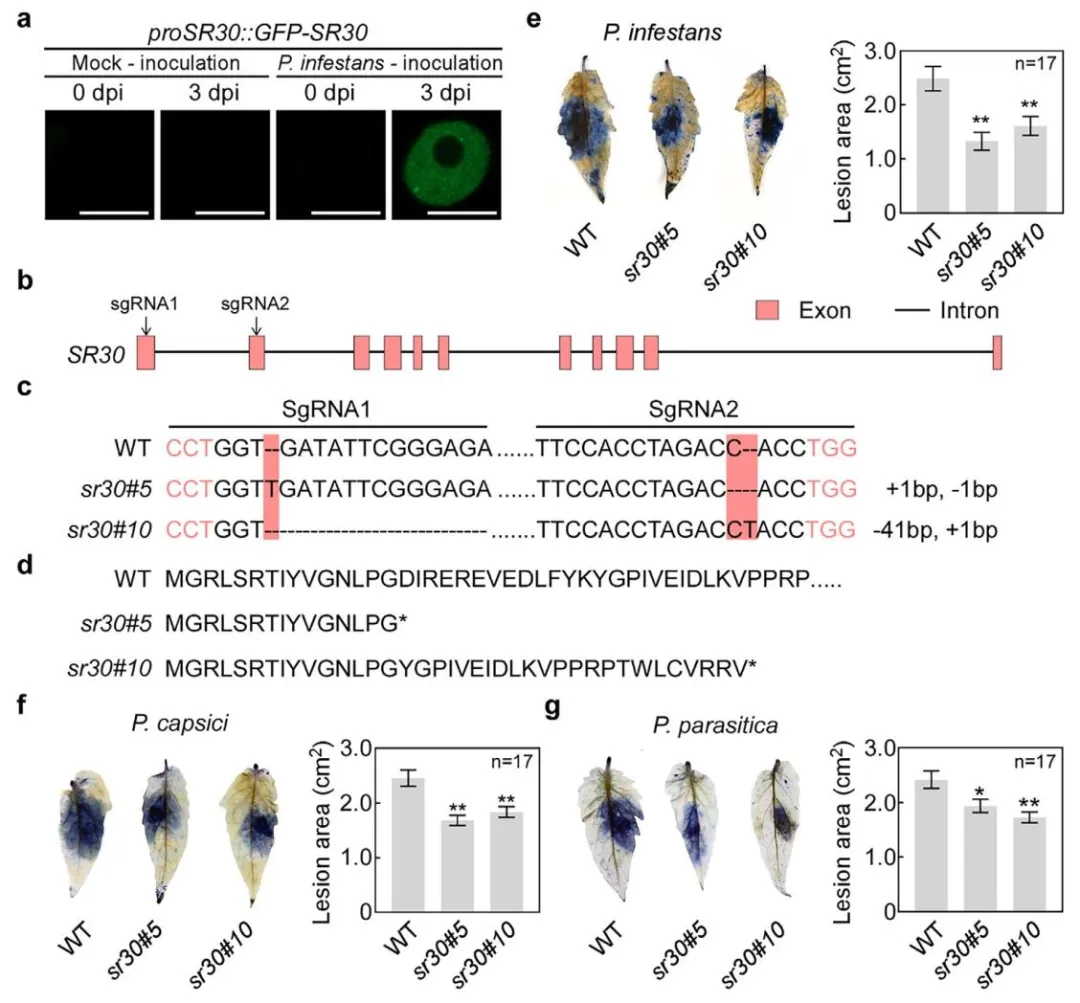
为了进一步验证敲除SR30的农业应用潜力,研究人员利用CRISPR/Cas9敲除SR30,构建突变体(sr30),随后接种多种病原体,观察抗性变化。结果发现sr30突变体对卵菌的抗性显著增强,但对真菌和线虫无影响。并且,与WT相比感染后,sr30中防御基因的功能性剪接比例显著升高(图6)。
图6. 敲除SR30增强番茄对三种卵菌病原体的抗性。
a)在致病疫霉菌(P. infestans)感染条件下,检测由SR30天然启动子驱动的GFP-SR30核凝聚物的形成。将携带proSR30::GFP-SR30载体的农杆菌浸润本氏烟草叶片,浸润区域在农杆菌浸润后1天接种模拟处理(Milli-Q水)或致病疫霉菌游动孢子。在接种模拟处理或致病疫霉菌后的0天和3天取样叶片组织,通过共聚焦显微镜观察并用ZEN软件拍照。比例尺为10微米。b)设计用于敲除SR30的两个小向导RNA(sgRNA)的示意图。箭头表示SR30基因中两个sgRNA的设计位置,红色方框代表外显子,黑色线条代表内含子。c)两个不同sr30突变体中SR30的基因组DNA(gDNA)序列。红色碱基表示两个原间隔相邻基序(PAM)的位置,红色区域通过序列比对显示两个sr30突变体与野生型(WT)的SR30 gDNA序列差异。d)sr30突变体和野生型番茄中SR30的蛋白质序列,星号表示终止密码子。e-g)图片显示,番茄中敲除SR30抑制了致病疫霉菌(e)、辣椒疫霉菌(f)和寄生疫霉菌(g)的病斑生长。对于致病疫霉菌,将sr30和野生型的离体叶片接种游动孢子,感染叶片用台盼蓝染色并在接种后4天(dpi)拍照。对于辣椒疫霉菌和寄生疫霉菌,将sr30和野生型的离体叶片接种菌丝块,感染叶片用台盼蓝染色并在接种后2天(dpi)拍照。柱状图显示三种疫霉菌感染的番茄(Micro-Tom)叶片的病斑面积。对于e-g,数据表示为平均值±标准误(SE,n=17),P值通过Student's t检验分析(*P<0.05,**P<0.01)。