背景介绍
免疫检查点抑制剂(ICIs)为晚期癌症带来了革命性治疗机遇,但临床数据显示,PD-1/PD-L1抗体单药治疗在大多数实体瘤中免疫应答率偏低,其疗效受限于单一的时空评估和靶向精度不足。近年来,金属在免疫调节中的作用备受关注——例如Mn²⁺可增强cGAS-STING通路敏感性,Zn²⁺参与胞质DNA识别。金属配合物具有更大的原子半径、更丰富的配位几何和更复杂的立体化学特征,使其在选择性识别生物大分子方面具备独特优势。然而,金属中心手性在精准靶向免疫检查点中的生物学作用,此前尚缺乏系统研究。
研究思路
针对这一科学问题,南京大学王秀秀教授、赵劲教授与魏炜教授团队合作,通过引入PD-L1结合基团BMS-8,构建了一对金属中心手性铱(III)纳米颗粒Δ-IrBMS8-NPs和Λ-IrBMS8-NPs,并进一步引入mPEG链使其直接自组装形成纳米颗粒,避免了纳米载体的使用。研究系统揭示了手性构型驱动的免疫检查点调控机制:Δ构型与PD-L1、Hapln1和Tagln具有更强的结合亲和力和更适配的空间结合构象(分子对接结合能分别为-3.96、-6.12和更高值),从而实现对PD-L1的立体选择性抑制。在PD-L1过表达的CT26结肠癌细胞中,Δ-IrBMS8-NPs在光照下的IC₅₀值低至0.131 μM,是Λ构型的19.5倍,光毒性指数高达763.4。更重要的是,Δ-IrBMS8-NPs通过特异性结合PD-L1,实现了129倍于无BMS-8单体Δ-Irph的肿瘤靶向富集。在CT26荷瘤小鼠模型中,Δ-IrBMS8-NPs联合光照处理实现了82%的肿瘤抑制率,显著激活了树突状细胞成熟(CD80⁺CD86⁺ DC比例达10.87%)、效应CD8⁺ T细胞浸润(CD8⁺IFN-γ⁺ T细胞达22.4%)和长期免疫记忆形成(记忆CD8⁺ T细胞达31.5%),同时有效抑制了调节性T细胞(Tregs)。相关内容以“Chirality-Driven Immune Checkpoint Modulation by Chiral Iridium(III) Nanoparticles”发表在《Journal of the American Chemical Society》!
图片解析
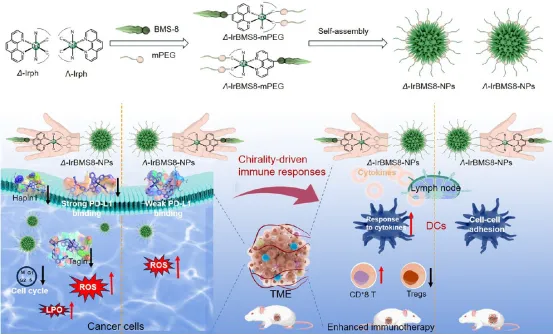
Scheme 1. 研究示意图:展示了手性铱(III)纳米颗粒Δ/Λ-IrBMS8-NPs的设计及其体外和体内手性驱动免疫检查点调控机制。Δ构型通过立体选择性识别PD-L1,增强肿瘤靶向、诱导多种细胞死亡(凋亡、坏死性凋亡、铁死亡)、激活树突状细胞和效应T细胞,最终重塑肿瘤微环境。
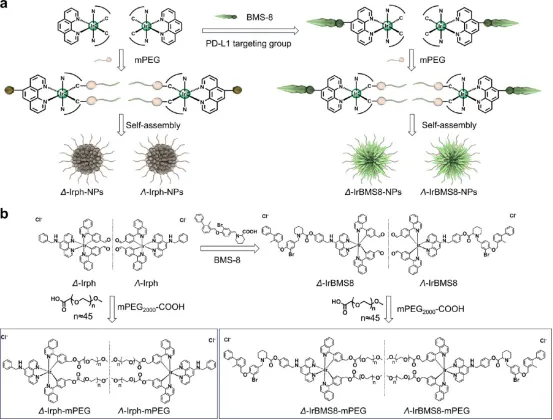 图1. 手性Ir(III)纳米颗粒的组装:(a) 手性免疫调节Ir(III)纳米颗粒设计示意图。(b) Δ/Λ-IrBMS8和Δ/Λ-Irph的合成路线。
图1. 手性Ir(III)纳米颗粒的组装:(a) 手性免疫调节Ir(III)纳米颗粒设计示意图。(b) Δ/Λ-IrBMS8和Δ/Λ-Irph的合成路线。
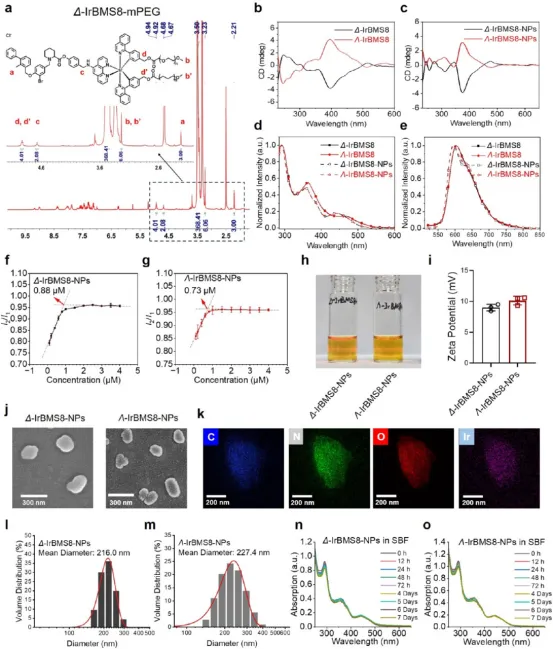
图2. 手性Ir纳米颗粒的表征:(a) Δ-IrBMS8-mPEG的¹H NMR谱,显示PEG链特征峰。(b-c) CD光谱显示Δ和Λ构型呈良好镜像关系,纳米颗粒保留单体手性。(d) UV-vis吸收光谱:<300 nm和~360 nm为配体中心跃迁,>400 nm为金属-配体电荷转移(MLCT)。(e) 光致发光光谱:纳米颗粒在~630 nm出现肩峰,归因于“疏水核-亲水壳”胶束结构形成。(f-g) CMC测定:Δ和Λ构型分别为0.88 μM和0.73 μM。(h) 廷德尔散射证实胶束形成。(i) Zeta电位约+10 mV。(j) SEM显示均匀球形纳米颗粒。(k) EDS证实C、N、O、Ir共定位。(l-m) DLS粒径:Δ构型~216.0 nm,Λ构型~227.4 nm。(n-o) SBF中7天UV-vis光谱稳定,证实生理稳定性。
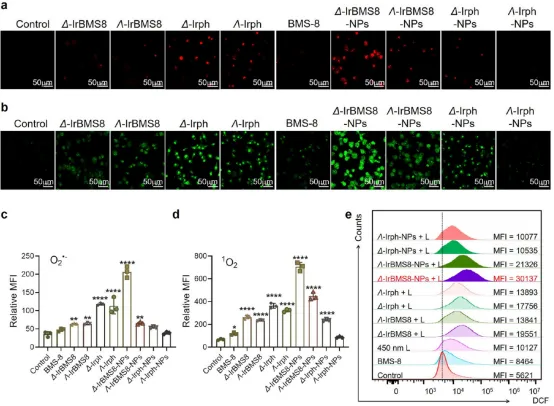
图3. 手性Ir纳米颗粒的抗癌疗效:(a-d) 共聚焦成像显示Δ-IrBMS8-NPs在CT26细胞中诱导最强的O₂⁻和¹O₂产生。(e) 流式细胞术证实Δ-IrBMS8-NPs诱导最强ROS积累。(光照条件:450 nm, 30 mW/cm², 10 min)
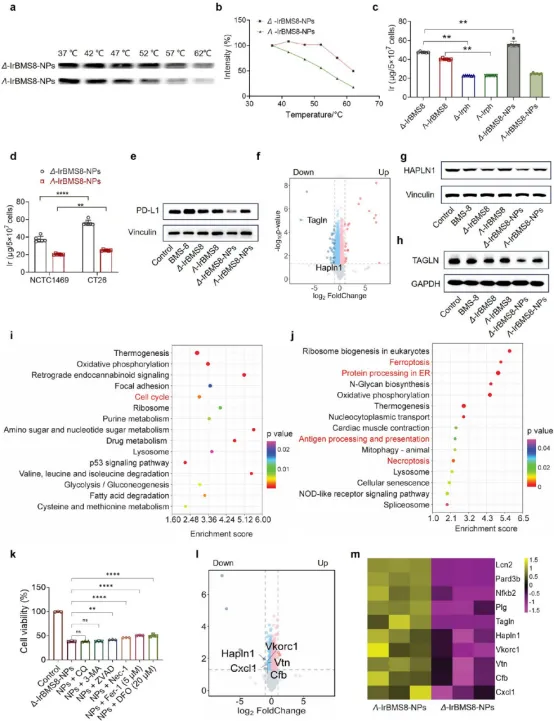
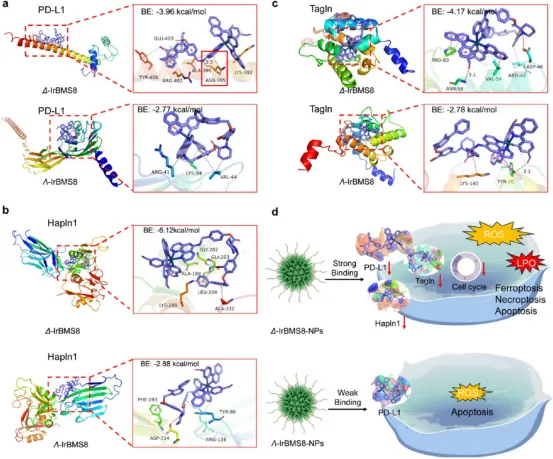
图4. 手性驱动抗癌活性的机制分析:(a-b) CETSA显示Δ-IrBMS8-NPs与PD-L1结合更稳定,热位移更显著。(c) ICP-MS显示CT26细胞中Δ-IrBMS8-NPs摄取量最高。(d) Δ-IrBMS8-NPs在正常细胞NCTC-1469中摄取极低,证实肿瘤选择性。(e) Western blot显示Δ-IrBMS8-NPs更有效抑制PD-L1表达。(f) Δ-IrBMS8-NPs vs 未处理细胞的蛋白质组火山图。(g-h) Western blot证实Δ-IrBMS8-NPs选择性抑制Hapln1和Tagln。(i-j) KEGG分析显示Δ构型下调氧化磷酸化和细胞周期通路,上调内质网蛋白加工和抗原加工呈递通路。(k) 细胞死亡抑制剂实验:Z-VAD(凋亡)、Nec-1(坏死性凋亡)、Fer-1(铁死亡)和DFO(铁螯合剂)均显著挽救Δ-IrBMS8-NPs诱导的细胞死亡,证实多种细胞死亡方式被激活。(l-m) Δ-IrBMS8-NPs vs Λ-IrBMS8-NPs的蛋白质组分析,Hapln1、Vkorc1、Vtn、Cfb、Cxcl1被Δ构型特异性抑制。
图5. 手性驱动抗癌活性的分子模拟机制:(a-c) 分子对接模拟显示Δ-IrBMS8与PD-L1、Hapln1、Tagln的结合能均高于Λ构型,且与PD-L1在ASN395位点形成显著氢键。(d) 机制示意图:Δ构型的立体选择性识别与结合驱动更强的抗癌活性。
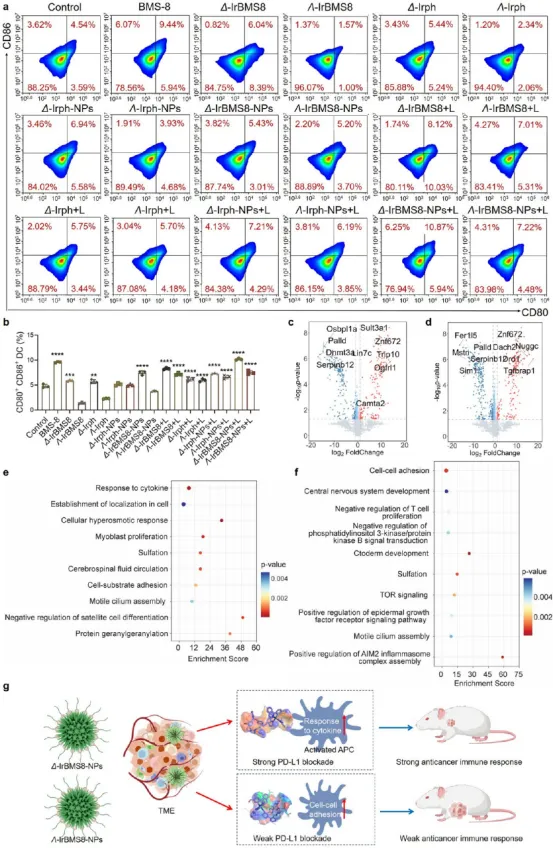
图6. 手性Ir(III)纳米颗粒在树突状细胞中的手性驱动机制分析:(a-b) 流式细胞术显示Δ-IrBMS8-NPs+L组肿瘤引流淋巴结中成熟DC(CD80⁺CD86⁺)比例达10.87%,为Λ构型的1.5倍。(c-d) 蛋白质组分析显示Δ构型处理DC中Ospblla、Palld、Dnmt3a、Lin7c显著下调,Sult3a1、Znf672、Trip10上调,富集于“response to cytokine pathway”。(e-f) Λ构型处理DC中Fer115、Mstn、Palld下调,Znf672、Nuggc、Dach2上调,富集于“cell-cell adhesion pathway”。(g) 示意图:Δ构型更有效激活DC成熟和细胞因子通路。
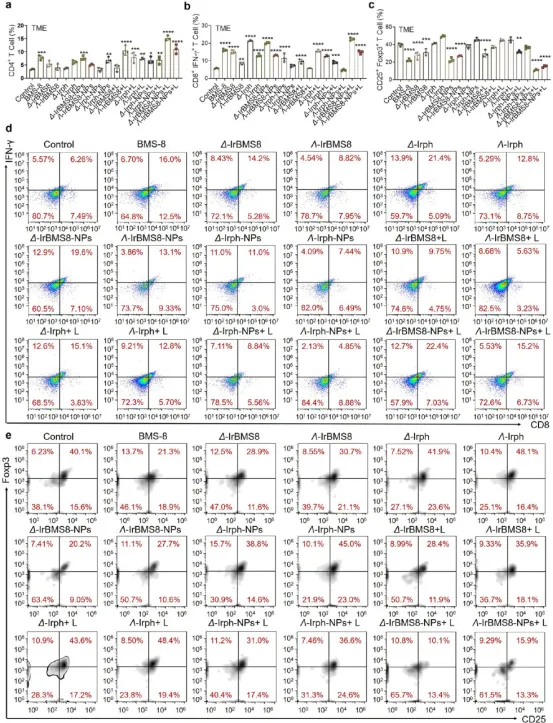
图7. 肿瘤微环境中免疫细胞分析:(a) Δ-IrBMS8-NPs+L组CD4⁺ T细胞比例达14.9%。(b,d) 效应CD8⁺ T细胞(CD8⁺IFN-γ⁺)在Δ构型组达22.4%,Λ构型组为15.2%。(c,e) 调节性T细胞(CD25⁺Foxp3⁺)在Δ构型组被最有效抑制。
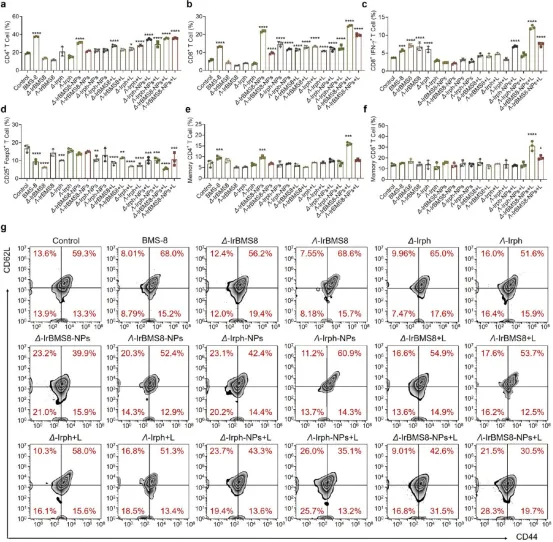
图8. 脾脏免疫细胞及免疫记忆分析:(a-d) Δ-IrBMS8-NPs+L组脾脏中CD4⁺、CD8⁺ T细胞及效应CD8⁺ T细胞比例最高,Tregs比例最低。(e-g) 记忆T细胞分析:Δ构型组记忆CD4⁺ T细胞达15.3%,记忆CD8⁺ T细胞达31.5%,显著高于Λ构型组(8.20%和19.7%)。
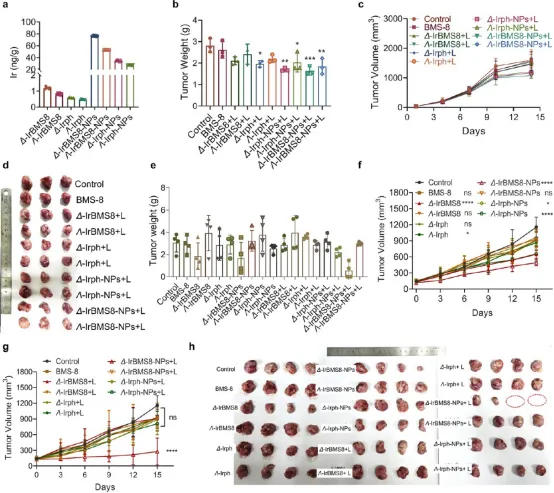
图9. 手性Ir(III)纳米颗粒的体内抗癌活性:(a) ICP-MS显示Δ-IrBMS8-NPs在肿瘤中的积累量是Δ-Irph单体的129倍,是Δ-IrBMS8单体的64倍,是Λ构型的1.44倍。(b-d) 免疫缺陷小鼠中,Δ-IrBMS8-NPs+L组肿瘤质量减少42%。(e-h) 免疫 Competent小鼠中,Δ-IrBMS8-NPs单独(无光照)抑制率约45%,联合光照后达82%;Λ构型无论有无光照效果均有限。
结论
本研究首次揭示了纳米尺度金属中心手性在精准靶向免疫检查点中的关键作用。通过引入PD-L1结合基团BMS-8并利用mPEG链诱导自组装,构建了一对手性铱(III)纳米颗粒Δ/Λ-IrBMS8-NPs。系统的手性驱动机制研究表明:Δ构型与PD-L1、Hapln1和Tagln具有更强的立体选择性结合亲和力,通过CETSA、蛋白质组学、分子对接等多维度验证。Δ-IrBMS8-NPs实现了129倍于无靶向单体Δ-Irph的肿瘤富集,在光照下IC₅₀低至0.131 μM(Λ构型的19.5倍),并诱导凋亡、坏死性凋亡和铁死亡等多种细胞死亡方式。在CT26荷瘤小鼠模型中,Δ-IrBMS8-NPs联合光照实现了82%的肿瘤抑制率,显著促进了树突状细胞成熟、效应CD8⁺ T细胞浸润和长期免疫记忆形成,同时有效抑制了调节性T细胞。该工作确立了Δ构型为增强抗癌免疫应答的最优立体化学结构,为开发高效手性免疫检查点抑制剂开辟了新方向。

「BioMed科技」关注生物医药×化学材料交叉前沿研究进展!交流、合作,请添加杨主编微信!

https://doi.org/10.1021/jacs.6c00020来源:BioMed科技声明:仅代表作者个人观点,作者水平有限,如有不科学之处,请在下方留言指正!