一、基本信息
文章题目:Bispericyclase guided discovery of nargenicin-type natural products
文章 DOI 号:https://doi.org/10.1016/j.apsb.2026.05.005
期刊名称:Acta Pharmaceutica Sinica B
通讯作者及工作单位:
- 戈惠明:南京大学生命科学学院,医药生物技术国家重点实验室,化学与生物医药创新中心,南京鼓楼医院神经内科 (State Key Laboratory of Pharmaceutical Biotechnology, Department of Neurology, Nanjing Drum Tower Hospital, Chemistry and Biomedicine Innovation Center, School of Life Sciences, Nanjing University)
- 张博:南京大学生命科学学院,医药生物技术国家重点实验室,化学与生物医药创新中心,南京鼓楼医院神经内科 (State Key Laboratory of Pharmaceutical Biotechnology, Department of Neurology, Nanjing Drum Tower Hospital, Chemistry and Biomedicine Innovation Center, School of Life Sciences, Nanjing University)
二、研究背景
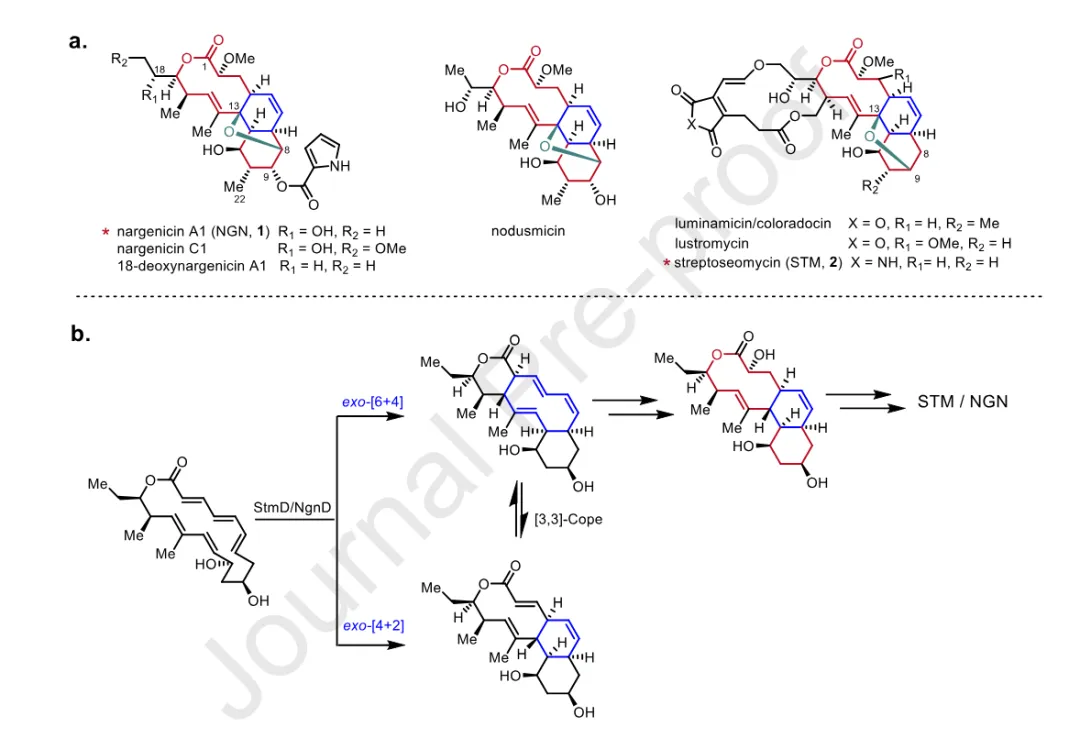
Nargenicin 类天然产物是一类结构独特的聚酮大环内酯抗生素,其结构特征在于一个含有醚桥的 10/6/6 三环核心骨架,该骨架由一个 cis-十氢萘 系统与一个含氧桥环共同构成。根据醚桥连接方式的不同,该类化合物可分为两个亚类:8,13-醚桥 亚类(以 nargenicin A1 为代表,C-9 位连有吡咯甲酸)和 9,13-醚桥 亚类(以 streptoseomycin 为代表,含马来酰亚胺或马来酸酐基团,形成 5/14/10/6/6 五环系统)。至今仅有 10 个 家族成员被报道,这与其生物合成潜力的规模极不相称。
Nargenicin A1 的作用机制已被阐明:其特异性抑制 DNA 聚合酶 III 的催化 α 亚基 DnaE,通过诱导 DNA 损伤发挥抗菌活性。Streptoseomycin 则对微需氧病原菌 Helicobacter pylori 表现出强效抑制活性。结构上的复杂性与生物活性的独特性,使得该类化合物的生物合成机制成为研究热点。本课题组前期的研究发现,NgnD 与 StmD 作为 双周环化酶 (bispericyclase),能够同时催化 [6+4] 与 [4+2] 周环反应,两者可通过 Cope 重排相互转化;其中 [6+4] 环加成的 6/10/6 加合物是真正的生物合成中间体,经羟化引发 [3,3]-Cope 重排与烯醇-酮互变异构后形成 10/6/6 三环骨架。
然而,该类化合物的完整生物合成途径,特别是聚酮骨架组装完成后的后续氧化、甲基化以及额外环系的形成步骤,仍 largely elusive。已报道的 BGCs 仅有 ngn(nargicin)与 stm(streptoseomycin)两个,极大地限制了组合生物合成与途径工程研究。因此,如何系统性地挖掘该类天然产物的生物合成潜力,发现新型同系物并阐明其修饰机制,是该领域的核心科学问题。
三、研究思路
本研究采用 保守双周环化酶引导的基因组挖掘 策略,系统性地探索 nargenicin 类天然产物的生物合成潜力与结构多样性。
作者以双周环化酶 StmD 为探针,在 NCBI nr 数据库及自有菌株库中进行同源性搜索,获得 452 个同源蛋白,通过系统发育分析构建了 nargenicin 家族的 BGC 蓝图。从中选取 8 株代表性菌株进行发酵筛选,发现 Nocardia tenerifensis NBRC 101015 产生三个新 nargenicin 同系物。针对其余 7 株未能产物的菌株,作者采用 CRISPR/Cas12a 辅助的 CAPTURE 技术 进行靶向捕获与异源表达,激活了其中的 sfa 与 cin 两个沉默 BGC。通过基因敲除、回补及体外酶活实验,解析了多个后修饰酶(P450 羟化酶、甲基转移酶)的功能。最后,对发现的新化合物进行抗菌活性评价与构效关系分析。该研究形成了“探针挖掘 → 蓝图构建 → 发酵筛选 → 沉默簇捕获与异源表达 → 功能验证 → 活性评价”的完整研究闭环。
四、研究方法
- 基因组挖掘与系统发育分析:以 StmD 为 query 序列,在 NCBI nr 数据库中进行 BLASTp 搜索(Expect threshold: 0.01),获得 452 个同源蛋白。使用 MEGA 等软件构建系统发育树,结合 antiSMASH 注释信息进行 BGC 分类与功能预测。
- CAPTURE 靶向捕获技术:采用 CRISPR/Cas12a 在体外精准酶切基因组 DNA 释放目标 BGC 片段,通过 T4 聚合酶 exon+ fill-in DNA assembly 与携带 loxP 位点的接收载体连接,经 Cre 重组酶介导的体内环化形成完整质粒,随后转化至 Streptomyces 异源宿主中进行表达。
- 异源表达:将 BAC 质粒 pHG0104(含 ngn 簇)及 CAPTURE 捕获的 sfa、cin 等 BGC 质粒,通过接合转移导入 S. albus J1074、S. lividans TK24、S. chartreusis 1018 及 S. coelicolor M1154 等宿主,在 R5A、SQ、F、No.1 等多种培养基中发酵,通过 HPLC-DAD 与 LC-MS 分析代谢产物。
- 靶向基因敲除:采用温度敏感型质粒 pKOV-kan 与 pDF25 介导的双交换同源重组系统,在 S. chartreusis 1018 中分别敲除 sfaP5、sfaO5 与 sfaM5 基因,通过 PCR 验证敲除成功与否,并比较突变株与野生型的代谢产物谱。
- 蛋白表达与体外酶活测定:将 ntfM1 克隆至 pET28a 载体,在 E. coli BL21(DE3) 中表达,经 Ni-NTA 亲和层析纯化。体外反应体系含 Tris-HCl (pH 7.4)、1 mmol/L 底物、1 mmol/L SAM、10 μmol/L NtfM1,30℃ 孵育 2 h 后通过 HPLC 分析产物。
- 化合物分离与结构鉴定:采用 MPLC(Biotage C18 柱)与半制备型 HPLC 分离纯化化合物。通过 HR-ESI-MS、1D NMR(¹H、¹³C)与 2D NMR(COSY、HMBC、NOE)确定平面结构;对化合物 10 尝试采用 Mosher 法测定绝对构型(未成功)。新化合物依据生物合成逻辑与光谱数据综合解析。
- 抗菌活性测定:采用微量肉汤稀释法测定化合物对 S. aureus 与 M. luteus 的 MIC 值,四环素为阳性对照。
五、实验设计及结果分析
(一) 基于 StmD 的基因组挖掘与 nargenicin 家族 BGC 蓝图构建
实验目的与设计逻辑
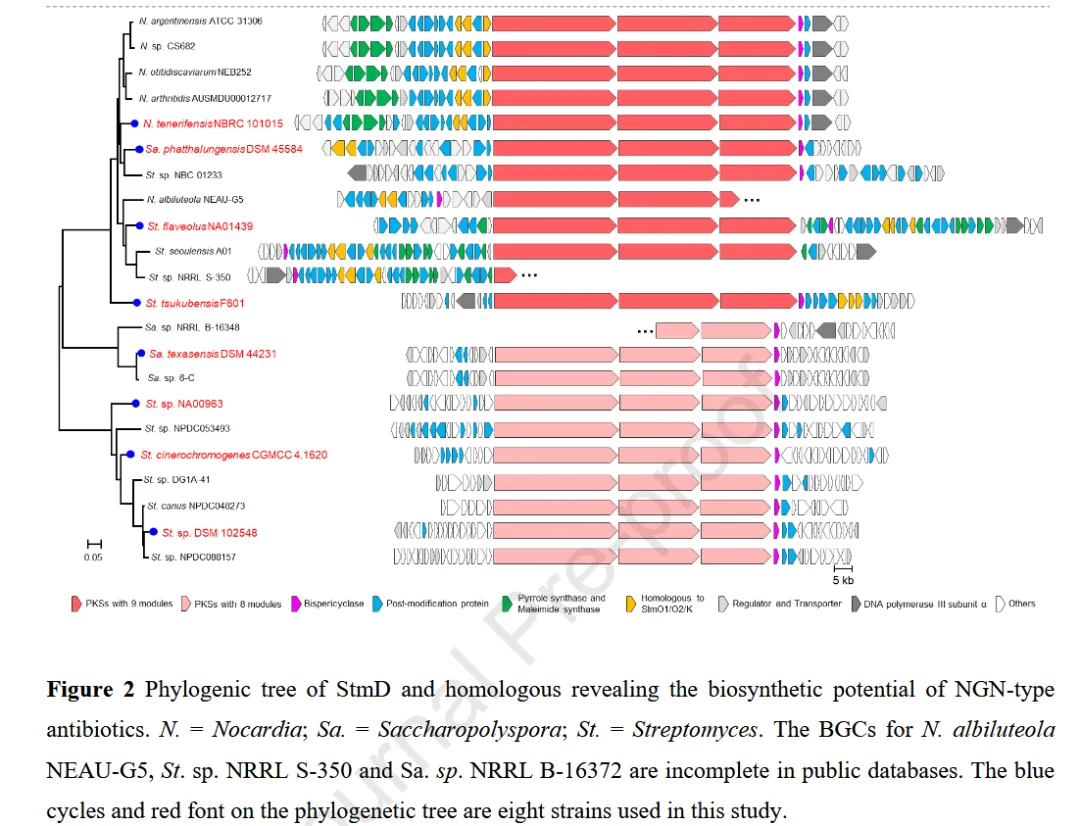
已知的 nargenicin 类 BGC 仅有 ngn 与 stm 两个,严重限制了该类天然产物的发现与工程化改造。为系统评估该类化合物的生物合成潜力,作者采用 保守双周环化酶 StmD(NTF2 家族 SnoaL2-like 聚酮环化酶,PF12680)作为探针,在 NCBI nr 数据库及自有菌株库中进行同源性搜索,构建系统发育树并建立 nargenicin 家族的 BGC 蓝图。
实验结果与机理解析
BLASTp 搜索共获得 452 个 StmD 同源蛋白,其中 291 个与次级代谢 BGC 无关被排除。图 S2 的系统发育分析显示,本研究鉴定的 StmD 同源物与已表征的其他 NTF2 家族酶(如 XiaE、SnoaL、IdmH)无显著序列相似性,提示该亚群经历了独立的进化轨迹。值得注意的是,StmD、NgnD 及前期表征的 101015D 与 59 个 额外同源物形成一个 distinct subclade,所有这些同源物均编码于 PKS BGCs 内部;而相邻 clade 则与 II 型芳香聚酮 的生物合成相关(图 S2、S3)。鉴于该 subclade 与 PKS BGCs 的专一性关联,作者将其归为 nargenicin 类天然产物的候选 BGC。
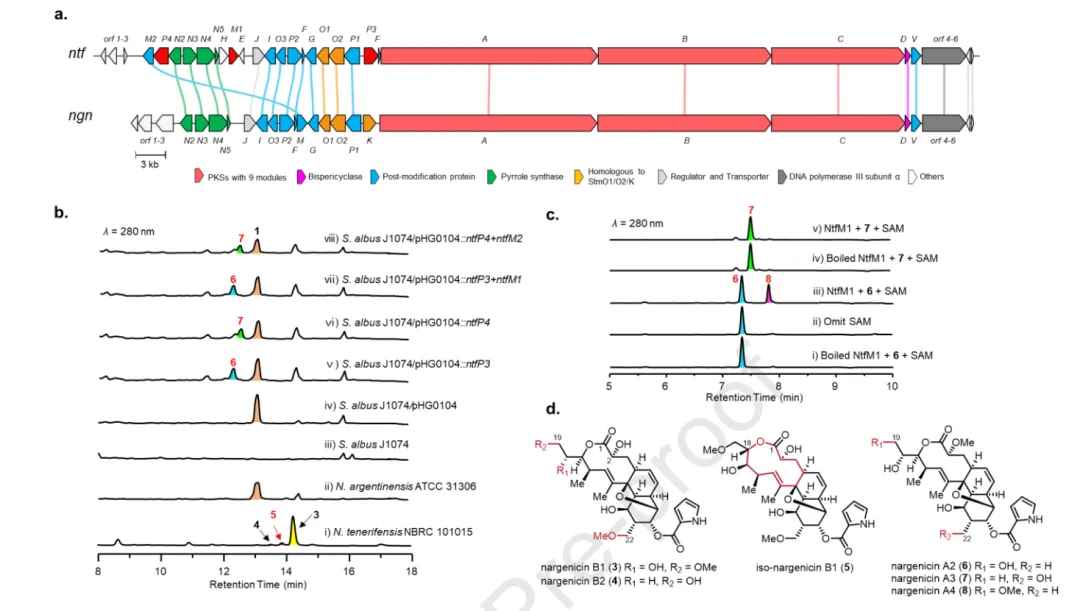
经过去冗余与低质量序列剔除后,作者建立了完整的 nargenicin 家族 BGC 蓝图(图 2)。在该蓝图中,4 个已报道的 nargenicin 产生菌株聚类在一起,而 STM 产生菌株与 Streptomyces sp. NRRL S-350 高度聚类且基因组织高度保守,提示后者为另一个 STM 生产者。根据 PKS 模块数目,这些 BGCs 可分为两类:9 模块型(如 ngn、stm)与 8 模块型(如 cin)。9 模块型 BGCs 编码多种后修饰酶,包括细胞色素 P450、Fe(II)/2OG 双加氧酶、甲基转移酶及自身抗性基因。图 2 中用蓝色圆圈标注了本研究选取的 8 株代表性菌株(其中 N. tenerifensis NBRC 101015 含 ntf 簇,S. flaveolus NA01439 含 sfa 簇,S. cinerochromogenes CGMCC 4.1620 含 cin 簇)。
(二) Nocardia tenerifensis 中新型 nargenicin 同系物的发现与生物合成解析
实验目的与设计逻辑
在上述 8 株候选菌株中,仅 N. tenerifensis NBRC 101015 在发酵筛选中产生 UV 吸收谱与 NGN A1 相似的化合物。为解析其结构并阐明生物合成机制,作者进行了大规模发酵、化合物分离与结构鉴定,并针对该菌株遗传操作困难的问题,构建了 ngn 簇的 BAC 质粒并在 S. albus 中进行异源表达,结合基因回补与体外酶活实验,鉴定了负责 C-19 与 C-22 羟化及甲基化的修饰酶。
实验结果与机理解析
图 3b 的 HPLC 分析显示,N. tenerifensis NBRC 101015 产生三个新峰 3-5。化合物 3(nargenicin B1)的 HR-ESI-MS 给出 [M+Na]⁺ 为 m/z 584.2470,分子式为 C₂₉H₃₉NO₁₀,比 NGN A1 ( 1 ) 多 46 Da(相当于两个甲氧基)。¹H NMR 显示两个甲氧基信号(δH 3.30 与 3.16),HMBC 分别相关至 C-19 (δC 74.9) 与 C-22 (δC 71.8);H-2 (δH 4.29) 无甲氧基相关信号,确定 C-2 为羟基而非甲氧基(图 S27)。化合物 4(nargenicin B2)分子式为 C₂₈H₃₇NO₉,仅含一个甲氧基(相关至 C-22),为 3 的 C-19-O-脱甲基类似物。化合物 5(iso-nargenicin B1)与 3 分子式相同,但关键 HMBC 相关 H-18 (δH5.04) 至 C-1 (δC 174.5) 提示为 11/6/6 三环骨架,作者提出其由 C-18 羟基对 C-1 酯羰基发生非酶促分子内酯交换形成(图 3d)。
图 3a 比较了 ntf 与 ngn 簇的基因组织:ntf 编码两个额外的 P450(NtfP3、NtfP4)与一个额外的甲基转移酶(NtfM1)。由于天然宿主遗传操作困难,作者构建了覆盖 ngn 簇的 BAC 质粒 pHG0104 并在 S. albus 中异源表达(图 S4)。将 ntfP3 或 ntfP4 单独导入该重组菌株后,分别产生两个新化合物 6 与 7(分子量均比 1 增加 16 Da),NMR 确证为 C-19 与 C-22 位甲基被羟化的衍生物(图 3d,表 S10-S11)。共表达 ntfM1 或 ntfM2 未检测到甲基化产物(图 3b)。
为确定甲基转移酶的功能,作者在 E. coli 中表达并纯化了 NtfM1(图 S5,NtfM2 未能获得可溶蛋白)。图 3c 与 图 S61 显示,在 SAM 存在下,NtfM1 特异性地转化 6(而非 7)生成分子量增加 14 Da 的产物 8。规模化酶反应后分离 8,NMR 确证为 C-19 位甲基化衍生物(图 3d,表 S12,图 S62-S68)。以上结果证明:NtfP3 与 NtfP4 分别催化 C-19 与 C-22 位羟化,NtfM1 催化 C-19 位 O-甲基化。鉴于 ntf 簇仅编码两个甲基转移酶,作者推测 NtfM2 负责 C-22 位 O-甲基化(图 S6)。
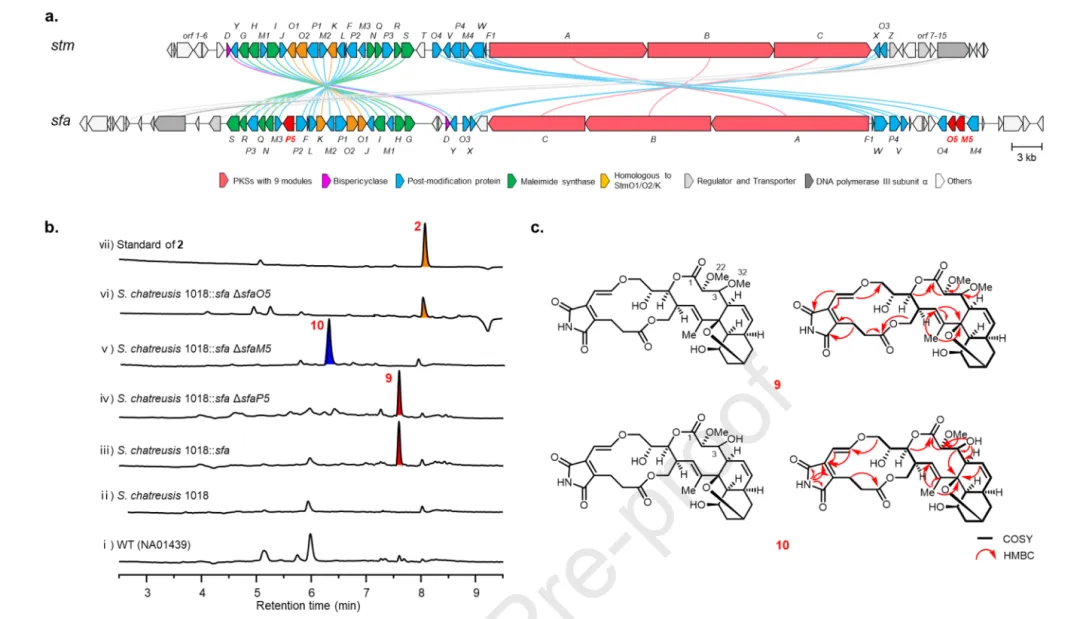
(三) 沉默 sfa BGC 的靶向捕获与 streptoflamycin 的发现
实验目的与设计逻辑
其余 7 株菌在多种发酵条件下均未检出 nargenicin 类产物,提示其 BGCs 在天然宿主中转录沉默。为激活这些隐秘 BGC,作者采用 CRISPR/Cas12a 辅助的 CAPTURE 技术 对目标 BGC 进行体外精准切割与克隆,随后导入 Streptomyces 异源宿主中进行表达。sfa 簇(来自 S. flaveolus NA01439,101 kb)是已鉴定同源簇中最大的一个,与 stm 簇高度相似但编码三个额外修饰酶(P450 SfaP5、Fe(II)/2OG 加氧酶 SfaO5 与甲基转移酶 SfaM5)。通过异源表达与靶向基因敲除,作者解析了这些修饰酶的功能。
实验结果与机理解析
图 4b panel iii 显示,sfa 簇在 S. chartreusis 1018 中异源表达后产生一个新代谢物 9,其 [M+H]⁺ 为 m/z 630.2541,UV 吸收与 STM ( 2 ) 一致且具有 360 nm 特征荧光(图 S8),提示含马来酰亚胺基团。HR-ESI-MS 确定分子式为 C₃₂H₃₉NO₁₂,比 2 多一个甲氧基。¹H NMR 显示两个甲氧基信号(δH 3.35 与 3.44),HMBC 分别相关至 C-2 (δC 87.5) 与 C-3 (δC 78.2),COSY 相关 H-2 (δH 3.74) 与 H-3 (δH 3.95) 确证 C-2 与 C-3 位各有一个甲氧基(图 S75)。该化合物被命名为 streptofulmycin (SFA, 9)。9 与已报道的 lustromycin 仅在马来酰亚胺与马来酸酐的差异上有别(后者 BGC 尚未报道)。
为解析三个额外修饰酶的功能,作者构建了 sfaP5、sfaO5 与 sfaM5 的框内缺失突变株(图 S7)。图 4b 显示:sfaP5 敲除突变株仍产生 9(panel iv),表明 SfaP5 在 SFA 生物合成中功能冗余;sfaO5 突变株不产生 9,而是积累一个与 2 保留时间和分子量相同的化合物(panel vi);sfaM5 突变株亦不产生 9,而是积累一个分子量比 9 少 14 Da 的化合物 10(panel v)。大规模发酵后分离鉴定:sfaO5 突变株产物为 2(streptoseomycin),sfaM5 突变株产物为 2 的 C-3 羟基衍生物 10(图 S78-S83,表 S14)。以上结果确证:SfaO5 催化 C-3 位区域选择性羟化,SfaM5 催化后续 O-甲基化(图 S9)。由于无法获得可溶的 SfaO5 与 SfaM5 蛋白,羟化与甲基化的精确时序尚不明确。图 4c 总结了 9 与 10 的结构及关键 HMBC 相关。
(四) 非经典 cis-AT PKS 催化的 endo-[6+4] 环加成产物 cinchromycin 的发现
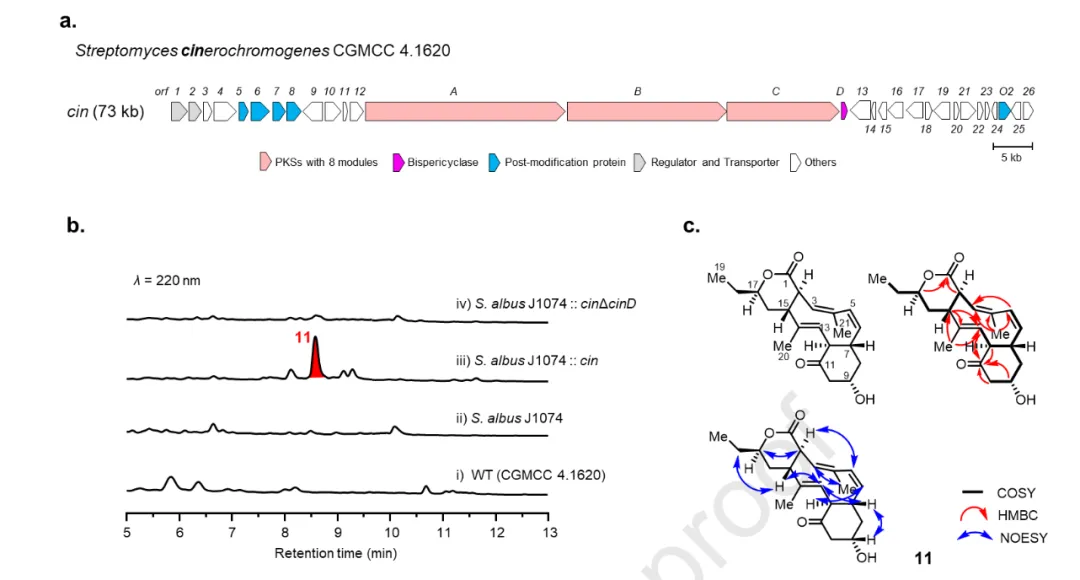
实验目的与设计逻辑
在 BGC 蓝图中,存在一个 8 模块型 的独特 phylogenetic clade,其 PKS 比 ngn/stm 少一个完整模块,模块 2 与/或模块 3 缺失 AT 结构域。作者选取 S. cinerochromogenes CGMCC 4.1620 的 cin 簇进行 CAPTURE 捕获与异源表达,旨在揭示该类非经典 PKS 的产物结构与环加成反应的立体化学特征。
实验结果与机理解析
图 5b 显示,cin 簇在 S. albus J1074 中异源表达后产生一个新代谢物 11,[M+Na]⁺ 为 m/z 367.1855,分子式 C₂₁H₂₈O₄。NMR 结构解析(图 5c,表 S16,图 S94-S99)揭示了一个 6/10/6 三环骨架,与已报道的 collinolactone A 和 rhizolutin 的 平面结构相同,但立体化学截然不同。关键 NOE 相关:H-21 (δ_H 1.84) 与 H-3 (δH 5.49) 相关,H-2 (δH 2.83) 与 H-5 (δH 6.01) 相关,支持连续的 Z,Z-构型二烯系统;H-7 (δH 2.13) 与 H-13 (δH 5.04) 相关,H-12 (δH 2.86) 与 H-6 (δH 5.25) 相关,表明 H-7 与 H-12 位于 6/10/6 系统的 opposite faces。这些 NOE 数据与生物合成逻辑共同确定了 11 为 endo-[6+4] 环加成加合物,与 STM 和 NGN 途径中表征的 exo-[6+4] 加合物(图 S1)形成鲜明对比。该化合物命名为 cinchromycin (CIN, 11),代表了一类 此前未被分离的天然产物。
图 5b 显示 cinD 基因敲除后 11 完全消失,证实 CinD 是催化该 endo-环加成的关键酶。值得注意的是,cin PKS 仅包含 7 个延伸模块,却 paradoxically 完成了 8 轮延伸(图 S10),且模块 2 与模块 3 缺失 AT 结构域——这些特征常见于 trans-AT PKS。为解析这一矛盾,作者构建了模块 2 与模块 3 的 KS 结构域系统发育树(图 S11),发现它们与 cis-AT PKS 聚类,而非 trans-AT 或 II 型 PKS。因此,cin 簇被归类为 cis-AT PKS。作者提出如下模型(图 S12):加载模块与模块 4 的 AT 结构域可能以 trans 方式 向模块 3 与模块 2 提供必需的延伸单元(甲基丙二酰-CoA 与丙二酰-CoA);模块 6 以迭代方式 进行第二轮链延伸以掺入额外的乙酸单元。
抗菌活性评价(表 S17)显示:C-22 位羟基化的 nargenicin A3 (7) 对 M. luteus 的 MIC 为 4 μg/mL,而缺乏 C-22 羟基的化合物 6 与 8 活性较弱(MIC 分别为 32 与 64 μg/mL),表明 C-22 羟化是增强抗菌活性的关键结构特征。
六、总体结论
本研究以保守双周环化酶 StmD 为探针,通过系统性基因组挖掘,构建了 nargenicin 类天然产物的 BGC 蓝图,将已知家族成员从 2 个 BGC 扩展至包含数十个同源簇的完整图谱。这是首次对该家族进行全局性生物合成潜力评估。
通过发酵筛选与 CAPTURE 介导的沉默簇激活,本研究发现了 3 类 新型 nargenicin 同系物:从 N. tenerifensis NBRC 101015 中鉴定了 nargenicin B1 ( 3 )、B2 ( 4 ) 与 iso-nargenicin B1 ( 5 ),并通过异源表达与体外酶活解析了 NtfP3/P4(P450 羟化酶) 与 NtfM1(甲基转移酶) 的功能;从 S. flaveolus NA01439 中激活了沉默 sfa 簇,发现了 streptofulmycin (9),并通过基因敲除确证 SfaO5(C-3 羟化酶) 与 SfaM5(C-3 甲基转移酶) 的功能;从 S. cinerochromogenes CGMCC 4.1620 中激活了 cin 簇,发现了 cinchromycin (11)——一个具有 endo-[6+4] 环加成 构型的 6/10/6 三环天然产物。
本研究最核心的机制发现 体现在两个方面:第一,CinD 催化 endo-[6+4] 环加成,与 StmD/NgnD 催化的 exo-环加成形成立体化学互补,拓展了人们对 周环化酶立体选择性的认识;第二,cin 簇的 PKS 虽为 cis-AT 类型,却表现出 trans-AT 的特征(模块缺失 AT、模块迭代),代表了 聚酮合酶编程的一种非经典范式。构效关系分析表明,C-22 位羟化是增强抗菌活性的关键修饰(化合物 7 对 M. luteus 的 MIC 为 4 μg/mL)。本研究为 nargenicin 类天然产物的发现、生物合成机制解析及工程化改造奠定了坚实的基础。
七、论文评价
优点与创新
本研究在 基因组挖掘策略 上具有高度创新性。不同于传统的基于核心 PKS 或骨架合成酶的挖掘,作者选择了 双周环化酶 StmD——一个在 nargenicin 类化合物生物合成中催化关键成环步骤的 功能标志酶——作为探针。该策略巧妙地将“结构决定功能”的逻辑反转为“功能特征引导发现”,显著提高了挖掘的特异性与效率。最终构建的 BGC 蓝图将已知家族从 2 个簇扩展到包含数十个同源簇的完整图谱,为其他天然产物家族的基因组挖掘提供了可推广的范式。
在 技术手段的综合性 上,本研究实现了从生物信息学(BLAST、系统发育、antiSMASH)到分子生物学(CAPTURE 靶向捕获、异源表达、基因敲除)再到生物化学(蛋白表达纯化、体外酶活)的全链条覆盖。特别值得指出的是,CAPTURE 技术 的应用使得 超过 100 kb 的大片段 BGC 能够被精准捕获并异源表达,克服了传统 cosmid 文库构建的繁琐与低效率问题,为激活大型沉默 BGC 提供了高效工具。
在 科学发现层面,cinchromycin (11) 的鉴定揭示了自然界中 endo-[6+4] 环加成 的存在,与已报道的 exo-环加成形成立体化学互补,拓展了周环化酶的催化多样性。同时,cin 簇 PKS 的非经典编程模式(cis-AT 骨架却表现出 trans-AT 特征)为聚酮合酶的结构-功能关系研究提出了新的问题。此外,构效关系分析 明确指出了 C-22 羟化 对抗菌活性的关键贡献,为后续药物化学优化提供了明确的修饰位点。
未来研究方向
尽管本研究成功鉴定了多个后修饰酶的功能,但部分酶的 体外重构 仍面临挑战。NtfM2 无法获得可溶蛋白,SfaO5 与 SfaM5 同样未能成功表达——这限制了对其催化机制(如底物谱、动力学参数、精确催化时序)的深入表征。未来可通过筛选不同的表达宿主(如 Streptomyces 自身表达系统)、融合标签(如 MBP、SUMO)或采用无细胞蛋白合成体系来解决蛋白可溶性表达问题。
cinchromycin (11) 中 endo-[6+4] 环加成的催化机制 尚待阐明。CinD 与 StmD/NgnD 的同源性比较与结构模拟可能揭示决定 exo/endo 选择性的关键残基。定点突变与体外环加成活性测定将是验证该机制的直接手段。此外,cin 簇 PKS 的 trans-AT 与迭代工作模型目前仍基于间接证据(KS 系统发育分析与碳原子计数),未来需要通过 体外 PKS 重构 或 模块置换实验 提供直接证据。
在生物活性方面,streptofulmycin ( 9 ) 与 C-3 羟基衍生物 10 的活性评价尚未完成。考虑到 9 含有两个额外的甲氧基(C-2 与 C-3),其抗菌活性谱与 potency 是否优于 STM ( 2 ) 值得系统评估。此外,化合物 5(iso-nargenicin B1)的 11/6/6 异常骨架是否由非酶促重排形成,还是存在专门的酶催化该反应,也是一个值得探索的问题。
八、关键问题及回答
Q1:本研究中作者以 StmD 作为探针进行基因组挖掘,而非使用核心 PKS 或更为保守的酮合成酶结构域。这一策略的优势是什么?为何 StmD 能够作为 nargenicin 家族 BGC 的“功能标志物”?
A:StmD(及同源的 NgnD)属于 NTF2 家族 SnoaL2-like 聚酮环化酶,其催化功能具有 高度特异性:StmD 同时催化 [6+4] 与 [4+2] 周环反应,这是 nargenicin/streptoseomycin 类天然产物 三环骨架形成的关键步骤。相比之下,核心 PKS 的 KS 结构域在聚酮合酶超家族中 高度保守,以 KS 为探针会产生大量假阳性(包括多种非目标聚酮的 BGC)。以 StmD 为探针的优势在于 功能-序列关联的特异性:只有负责合成含 [6+4]/[4+2] 环加成产物的 BGC 才会编码此类双周环化酶。因此,该策略大幅提高了挖掘的“信噪比”。本研究结果也验证了这一策略的有效性:452 个 同源蛋白中,与 PKS BGC 关联的 161 个 均被归入 nargenicin 家族,无假阳性簇被误纳入。此外,该策略还可识别“功能失活”的 BGC——例如,若某同源簇编码的 StmD 同源物发生了关键催化残基的突变,作者可据此推断该簇可能为“伪基因簇”。这一方法学为其他含有 特征性周环化酶 的天然产物家族(如含 Diels-Alderase 的家族)的基因组挖掘提供了可推广的范式。
Q2:cin 簇的 PKS 仅编码 7 个延伸模块,却完成了 8 轮延伸,且模块 2 与模块 3 缺失 AT 结构域。作者将其归类为 cis-AT PKS 并提出了 trans-AT 与迭代的工作模型。请从聚酮合酶的结构-功能关系角度,详细阐述这一非经典编程模式的可能分子机制。
A:经典的 cis-AT PKS 每个模块均包含一个完整的 AT 结构域,负责选择并将延伸单元装载至相邻的 ACP 上;而 trans-AT PKS 则缺乏 AT 结构域,由游离的 AT 酶以 trans 方式提供延伸单元。cin 簇呈现出“混合”特征:系统发育分析(图 S11)显示其 KS 结构域与 cis-AT PKS 聚类,但模块 2 与模块 3 缺失 AT。作者提出的模型如下:模块 4 的 AT 结构域(仍保留)可能以 trans 方式 向 模块 3 提供甲基丙二酰-CoA 延伸单元;加载模块的 AT 结构域 可能以 trans 方式 向 模块 2 提供丙二酰-CoA 延伸单元。这种“模块间 trans-AT 互补”在天然产物生物合成中虽不常见但已有先例。关于 模块迭代:聚酮链延伸至模块 6 后,通常应进入硫酯酶结构域环化释放。然而,cin 簇的终产物 11 含有 21 个碳,与 7 个模块的单轮延伸(1 个起始单元 + 7 个延伸单元 = 8 个二碳单元,共 16 个碳)不符。因此,模块 6 必须迭代使用:在第一轮延伸中,模块 6 催化添加一个延伸单元;随后,聚酮链 不进入 TE 结构域,而是返回至模块 6(或模块 5-6 的接头区域)进行第二轮延伸,添加第二个乙酸单元。KS6 结构域可能具有 接受不同链长底物 的灵活性,或其活性位点口袋能够 accommodate 更长的聚酮链。体外重构 KS6-AT6-ACP6 三联体结构域并测定其对不同链长底物的催化活性,将是验证该迭代模型的最直接手段。该非经典 PKS 编程模式不仅拓展了聚酮合酶的结构-功能认知,也为通过模块重排产生“非天然”聚酮化合物提供了新的工程化策略。
Q3:化合物 10(3-羟基 streptoseomycin 衍生物)的绝对构型未能通过 Mosher 法确定——衍生化仅得到 C-11 与 C-18 位的双 MTPA 酯,C-3 位未发生酯化。请从化学结构的角度分析导致这一失败的可能原因,并建议替代的绝对构型确定方法。
A:Mosher 法(α-甲氧基-α-三氟甲基苯乙酸酯法)测定仲醇绝对构型的前提是目标羟基能够与 MTPA 氯(或酸酐)发生 高效酯化反应。化合物 10 中 C-3 羟基未能酯化的可能原因包括:第一,空间位阻:C-3 位可能处于高度拥挤的化学微环境中——相邻的 C-2 位甲氧基与 C-4 位取代基可能形成“口袋”,阻碍 MTPA 氯的接近;第二,氢键网络:C-3 羟基可能与邻近的羰基氧(如 C-1 内酯羰基)形成分子内氢键,降低其亲核性;第三,电子效应:C-3 位的电子云密度可能因共轭体系的影响而降低,不利于亲核取代反应。替代方法 包括:第一,改良的 Mosher 法:尝试使用更活泼的衍生化试剂如 MTPA 酸酐(而非 MTPA 氯),或在更强碱(如 DMAP)存在下进行反应;第二,CD 激子手性法:若能将 C-3 羟基与对甲氧基苯甲酸(p-methoxybenzoic acid)或萘甲酸等发色团衍生化,通过比较衍生物的 CD 谱图与激子耦合模型可确定绝对构型;第三,化学转化结合 NMR:将 10 氧化为 C-3 酮,再通过立体选择性还原获得两个非对映异构体,比较其 NMR 化学位移差异推测原始构型;第四,TDDFT-ECD 计算:直接测定 10 的实验 ECD 谱,计算其 (3R) 与 (3S) 异构体的理论 ECD 谱进行比对——该方法无需衍生化,特别适用于 微量样品 或 衍生化困难 的化合物,本研究在解析化合物 4(深海真菌论文)时已成功应用该方法。鉴于 10 的量可能有限(从突变株发酵获得),TDDFT-ECD 计算 可能是最切实可行的选择。
SynBioPath — 合成生物学研究平台,收录 300+ 精选科研工具与数据库资源,涵盖 AI 搜索、文献管理、基因/蛋白分析、生物合成与代谢、天然产物研究等全流程工具链。
平台还提供常用科研软件的入门与进阶教程,以及天然产物、生物合成与酶工程等领域的基础学习资料,帮助研究者快速上手。
欢迎访问:www.synbiopath.com