药物降脂致心缺陷:为验证人群发现的因果关系,研究人员采用两种叶酸代谢干扰药物处理怀孕母鼠:甲氨蝶呤(抑制二氢叶酸还原酶)和磺胺嘧啶(抑制肠道菌群叶酸合成)【图2A】。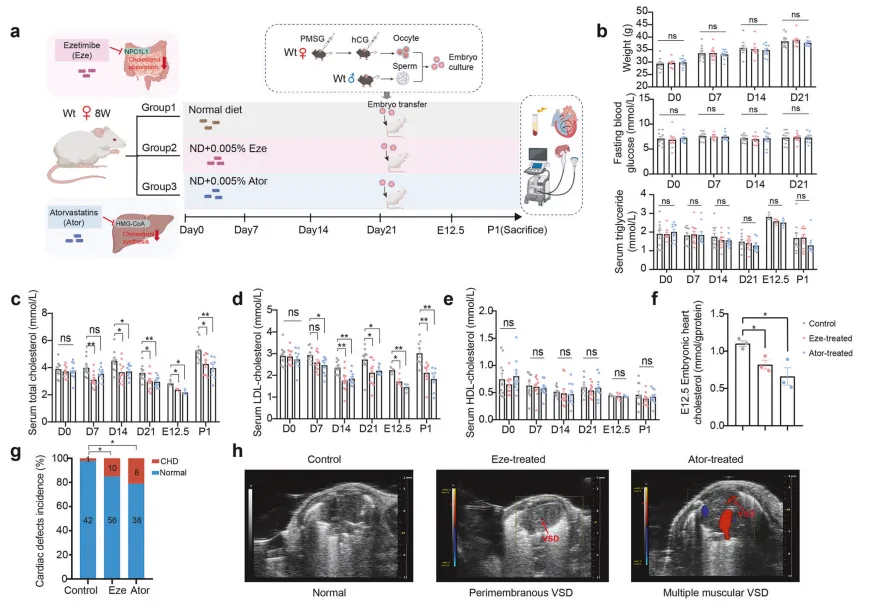 结果显示,药物处理成功降低了孕鼠血清及胚胎神经管的叶酸水平【图2C-F】,并直接导致子代神经管缺陷发生率显著升高:对照组仅为1.8%,甲氨蝶呤组升至16.5%,磺胺嘧啶组达14.2%【图2G】,证实孕期母体叶酸短期缺乏即可严重干扰胚胎神经管闭合过程。
结果显示,药物处理成功降低了孕鼠血清及胚胎神经管的叶酸水平【图2C-F】,并直接导致子代神经管缺陷发生率显著升高:对照组仅为1.8%,甲氨蝶呤组升至16.5%,磺胺嘧啶组达14.2%【图2G】,证实孕期母体叶酸短期缺乏即可严重干扰胚胎神经管闭合过程。
对86例神经管缺陷患儿的全基因组测序显示,叶酸代谢关键基因MTHFR的功能性突变(C677T)与疾病高度相关。
该突变在人群中携带率较高,在两个独立出生队列验证中均显示,携带者神经管缺陷风险为正常人群的2.8~3.5倍。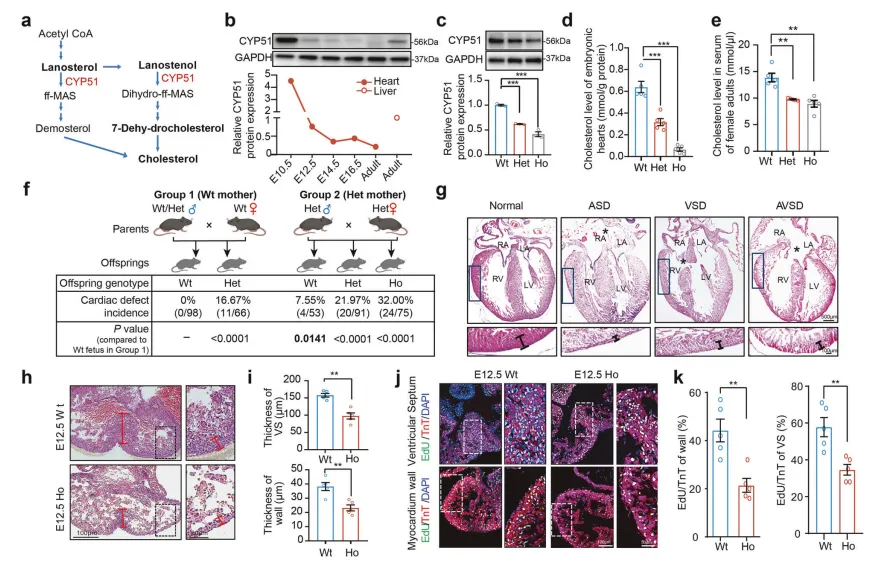
构建携带同源突变的Mthfr基因敲入小鼠模型证实,该突变导致MTHFR酶活性显著下降【图3C】,纯合突变小鼠胚胎神经管及胚胎组织中叶酸水平降低约70%【图3D】,28%的新生纯合突变小鼠出现脊柱裂、无脑畸形等神经管缺陷表型【图3F-H】。
核心机制在于突变体神经上皮细胞增殖能力显著受损,导致神经褶闭合延迟、融合失败【图3J, K】。
单细胞测序分析发现,突变体神经上皮细胞中胚胎发育关键通路Wnt/β-catenin的活性显著抑制【图4A, B】,该通路的信号传导高度依赖叶酸介导的DNA甲基化修饰: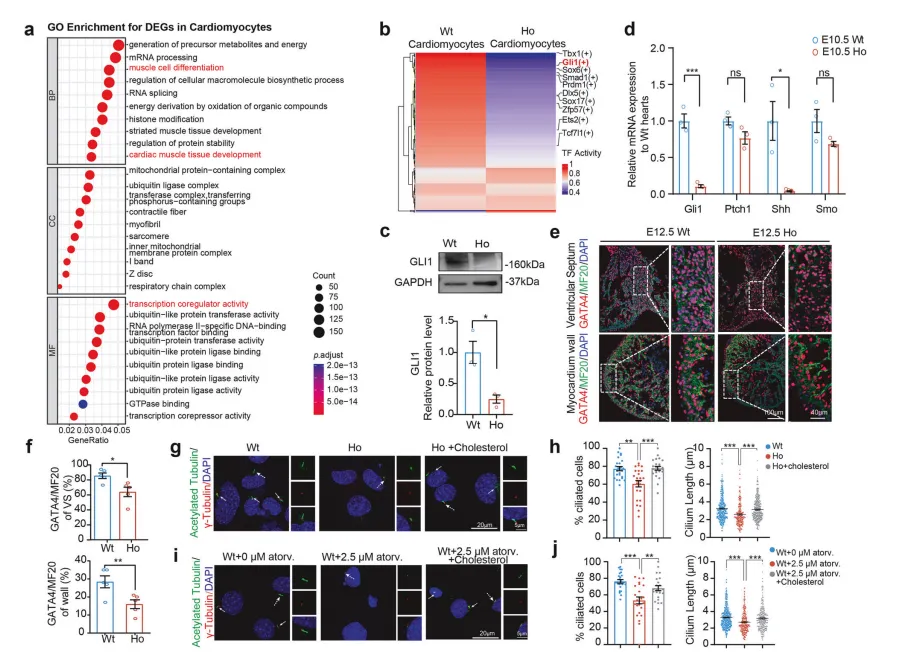
直接抑制通路核心蛋白:突变体中Wnt/β-catenin通路核心效应蛋白β-catenin的核内积累显著减少【图4C】;
破坏信号接收微环境:神经上皮细胞顶端的肌动蛋白骨架是维持Wnt信号极性的结构基础,叶酸缺乏导致骨架紊乱、极性丧失,信号传导效率大幅下降【图4G-J】。
β-catenin可直接结合神经管闭合关键转录因子Pax3的启动子调控其表达,最终形成“叶酸缺乏→骨架紊乱→Wnt信号减弱→Pax3表达下降→神经管闭合缺陷”的完整致病链条【图4E, F】。
“挽救实验”显示,给携带Mthfrᶜ⁶⁷⁷ᵀ突变的怀孕母鼠补充高剂量5-甲基四氢叶酸后,纯合突变子代小鼠的神经管缺陷发病率从28.3%大幅降至6.9%【图5C】。
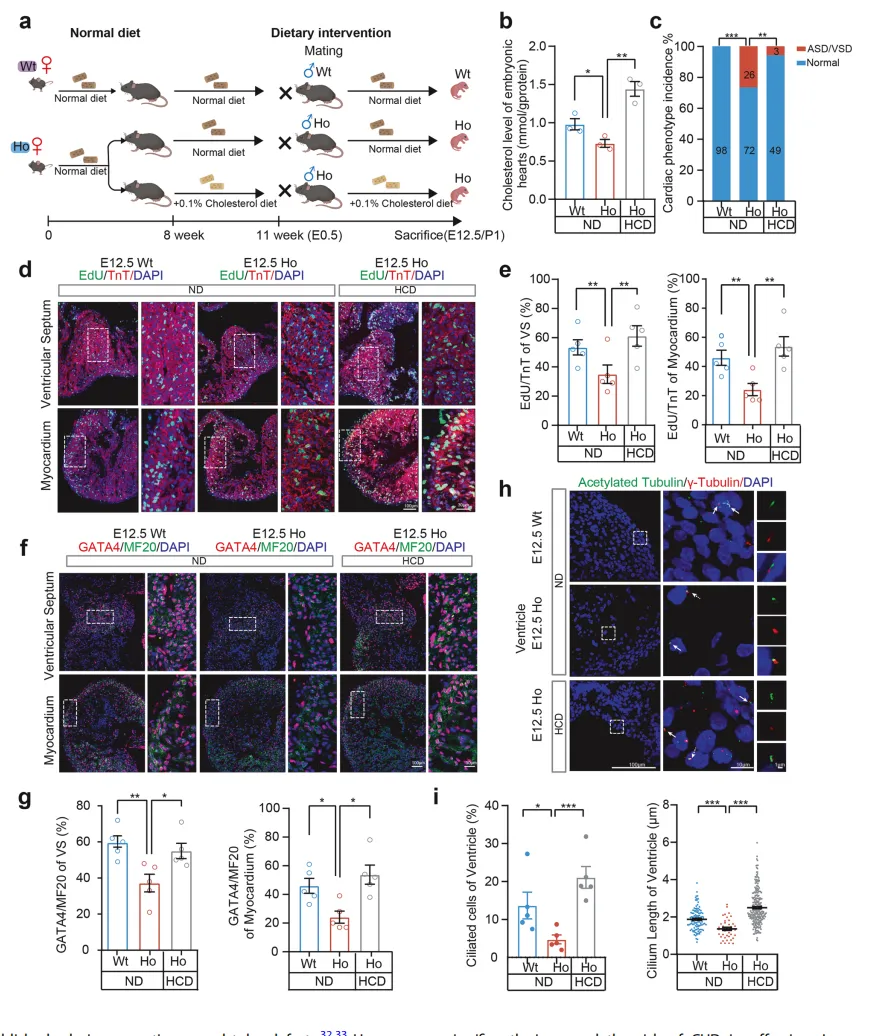 补充5-甲基四氢叶酸不仅恢复了胚胎神经管的叶酸水平【图5B】,还同步提升了神经上皮细胞的增殖活性【图5D, E】、恢复了Pax3和β-catenin的表达【图5F, G】,并修复了肌动蛋白骨架的极性排列【图5H, I】。
补充5-甲基四氢叶酸不仅恢复了胚胎神经管的叶酸水平【图5B】,还同步提升了神经上皮细胞的增殖活性【图5D, E】、恢复了Pax3和β-catenin的表达【图5F, G】,并修复了肌动蛋白骨架的极性排列【图5H, I】。




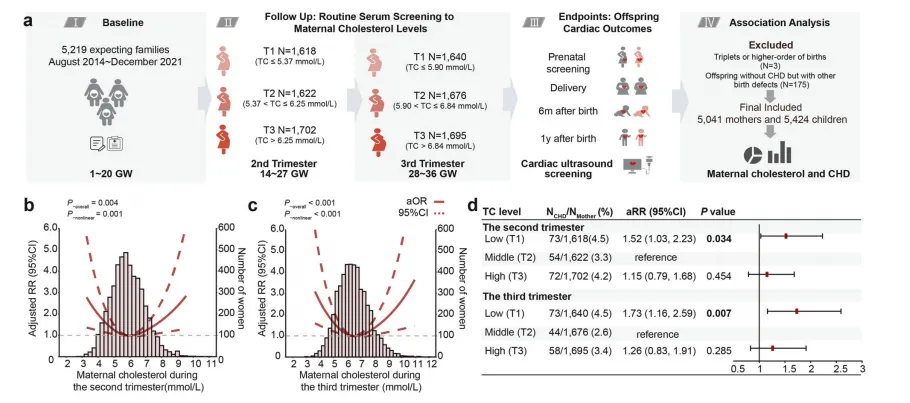 其中叶酸缺乏群体风险尤为突出:与叶酸水平处于中间区间(T2)的孕妇相比,孕早期叶酸最低组(T1)子代神经管缺陷风险升高2.1倍,孕中期该风险进一步升至3.4倍【图1D】。这是首次在中国大样本队列中证实孕早期叶酸缺乏是神经管缺陷的独立危险因素。
其中叶酸缺乏群体风险尤为突出:与叶酸水平处于中间区间(T2)的孕妇相比,孕早期叶酸最低组(T1)子代神经管缺陷风险升高2.1倍,孕中期该风险进一步升至3.4倍【图1D】。这是首次在中国大样本队列中证实孕早期叶酸缺乏是神经管缺陷的独立危险因素。
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
