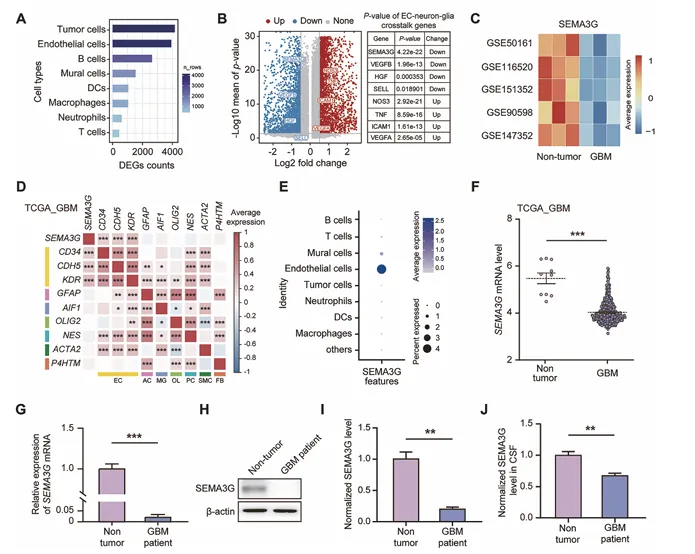
CellChat分析显示,肿瘤细胞是SEMA3G的主要受体细胞。AP标记的SEMA3G直接结合于GBM肿瘤组织,且被NRP2中和抗体阻断(图2)。
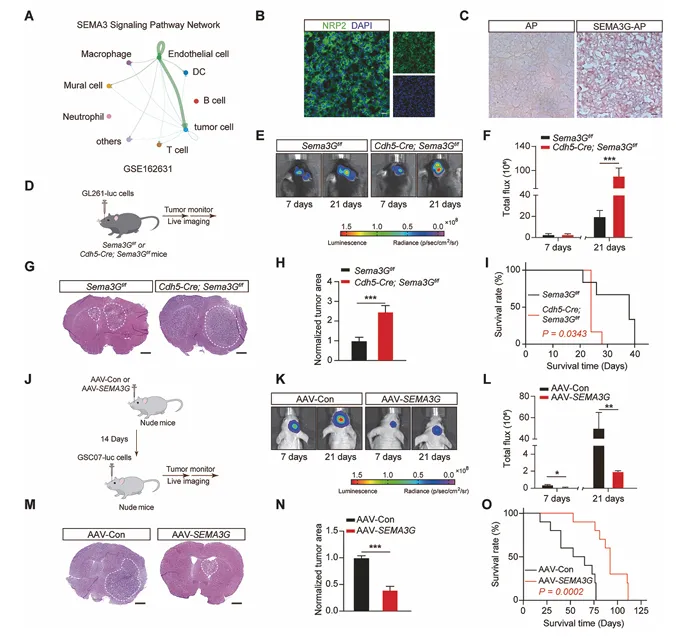
图2. ECs来源的SEMA3G抑制体内GBM生长
ECs特异性Sema3g敲除小鼠中,GL261细胞原位移植后肿瘤生长加速,生存期缩短。相反,SEMA3G过表达或重组SEMA3G蛋白处理显著抑制肿瘤生长,延长生存期。
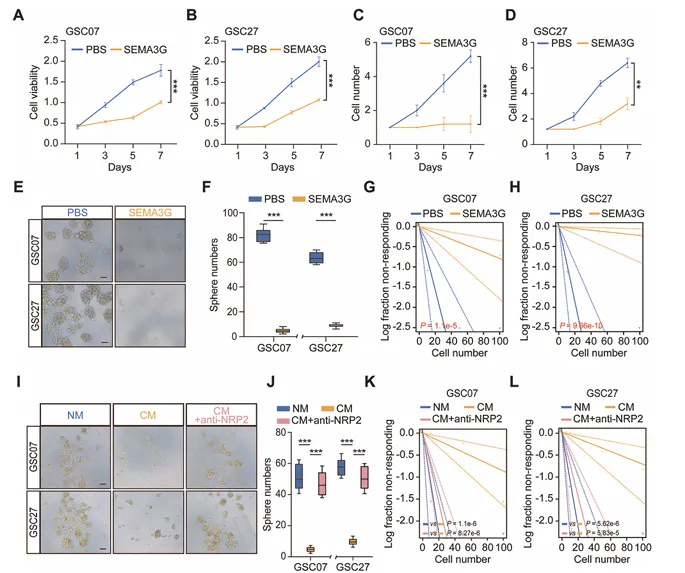
重组SEMA3G蛋白处理GSC07和GSC27细胞,剂量和时间依赖性降低细胞活力和自我更新能力,抑制球体形成。极限稀释实验证实SEMA3G损害GSC干性(图3)。
SEMA3G过表达HUVEC的条件培养基同样抑制GSC干性,且该效应被NRP2中和抗体逆转,证实SEMA3G介导ECs-GSC互作。
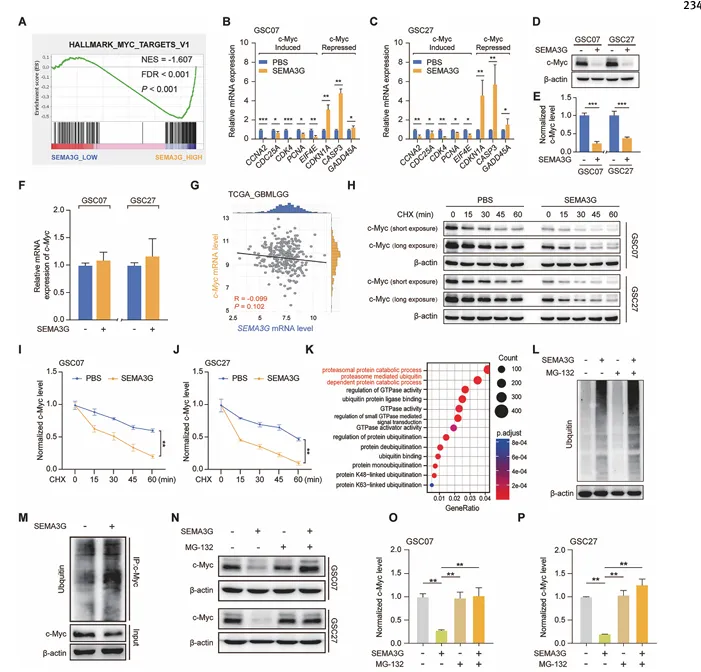
GSEA分析显示,SEMA3G低表达患者富集c-Myc靶基因。SEMA3G处理GSC细胞后,c-Myc蛋白水平降低,而mRNA水平不变。CHX chase实验显示SEMA3G缩短c-Myc半衰期(图4)。
图4. SEMA3G降低GSC中c-Myc蛋白稳定性
GO分析提示蛋白酶体通路参与。SEMA3G增加c-Myc泛素化水平,MG-132可逆转SEMA3G诱导的c-Myc下调,证实SEMA3G通过泛素-蛋白酶体途径促进c-Myc降解。
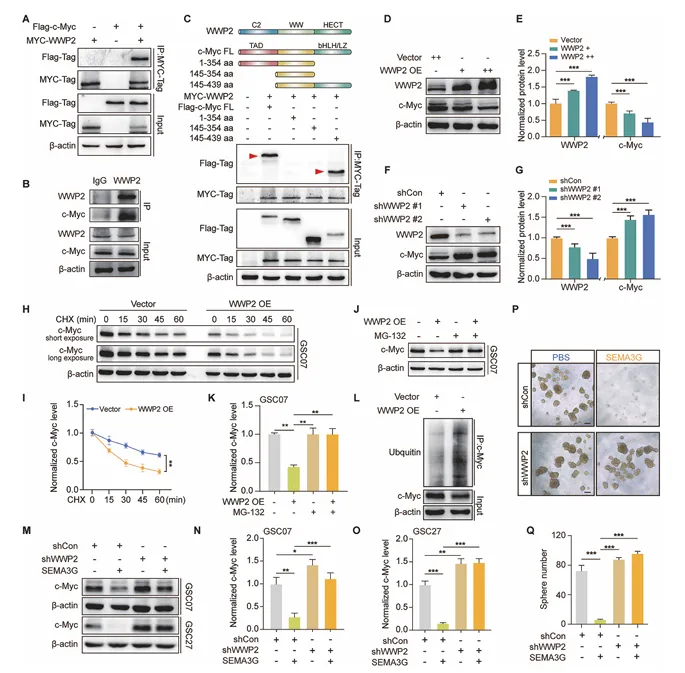
通过Ubiqowser数据库预测和STRING分析,筛选出与c-Myc相互作用的候选E3连接酶,其中WWP2与c-Myc结合最强。Co-IP证实内源性WWP2与c-Myc互作,c-Myc的bHLH/LZ结构域是关键结合区域(图5)。
图5. SEMA3G以WWP2依赖性方式促进c-Myc降解
WWP2过表达降低c-Myc蛋白水平、缩短其半衰期、增加其泛素化;WWP2敲低则相反。更重要的是,WWP2敲低可逆转SEMA3G诱导的c-Myc降解和球体形成抑制。
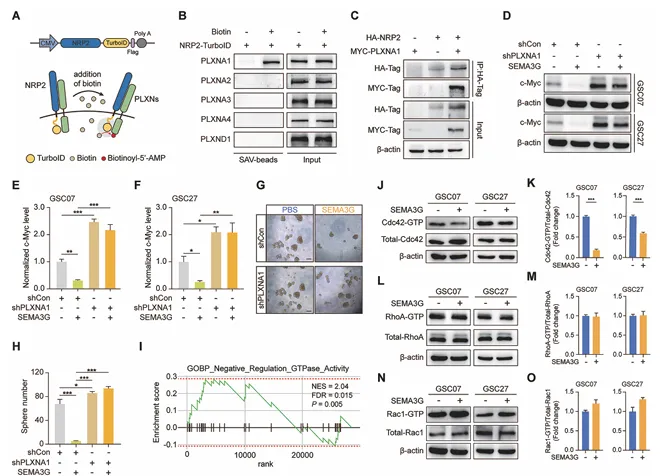
SEMA3G通过NRP2/PLXNA1失活Cdc42
TurboID邻近标记鉴定PLXNA1为NRP2的共受体。Co-IP证实NRP2与PLXNA1互作。PLXNA1敲低可逆转SEMA3G诱导的c-Myc降解和球体形成抑制。GO分析提示SEMA3G与GTPase活性负调控相关(图6)。
图6. SEMA3G通过NRP2/PLXNA1失活Cdc42]
SEMA3G处理显著降低Cdc42-GTP水平,而对RhoA和Rac1活性无显著影响,提示SEMA3G特异性失活Cdc42。












 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
