科研 | 南京中医药大:甘草次酸通过Thbs1/PI3K-Akt/p53通路调节肠道菌群及其代谢物进而改善胃黏膜损伤
- 2026-06-14 09:05:30
 点击蓝字“代谢组metabolome”,轻松关注不迷路
点击蓝字“代谢组metabolome”,轻松关注不迷路

生科云网址:https://www.bioincloud.tech/
编译:微科盟 草重木雪,编辑:微科盟X、江舜尧。
微科盟原创微文,欢迎转发转载。
肠道菌群的菌群失调在胃黏膜损伤(GMI)的发展中起着关键作用。甘草次酸(GA)是一种生物活性三萜类化合物,大量存在于甘草根中。尽管GA在减轻GMI方面的潜力已得到认可,但其确切机制仍然不清楚,特别是关于肠道微生物群的作用。本研究旨在探讨GA在预防大鼠盐酸(HCl)/乙醇诱导的GMI中的保护作用和机制。H&E染色揭示形态和结构证明了GA对胃黏膜的保护作用。研究者利用粪便微生物群移植,GA通过肠道微生物群依赖性机制显著减轻GMI大鼠的氧化损伤、炎症和细胞凋亡相关基因的表达。16S rRNA测序和代谢组学分析显示,GA改善了HCl/乙醇触发的肠道菌群失调和鞘脂、精氨酸和色氨酸代谢的失衡;促进肠道微生物群中婴儿双歧杆菌(B. infantis)的丰度;改善了与损伤相关的代谢紊乱。此外,其作用机制与抑制Thbs1/PI3K-Akt/p53信号通路有关。因此,GA通过调节Thbs1/PI3K-Akt/p53通路为GMI提供治疗潜力,从而减轻与肠道菌群失衡相关的炎症反应。
亮点:
1. 甘草次酸(GA)通过调节肠道菌群减轻胃黏膜损伤。
2. GA可以促进精氨酸和1-磷酸鞘氨醇的产生;
3. GA可以富集婴儿双歧杆菌并调节鞘脂、精氨酸和色氨酸代谢;
4. GA通过Thbs1/PI3K-Akt/p53通路减轻胃黏膜损伤。
论文ID
原名:Glycyrrhetinic acid ameliorates gastric mucosal injury by modulatinggut microbiota and its metabolites via Thbs1/PI3K-Akt/p53 pathway
译名:甘草次酸通过Thbs1/PI3K-Akt/p53通路调节肠道微生物及其代谢物,从而改善胃黏膜损伤
期刊:Phytomedicine
IF:8.3
发表时间:2025.4
通讯作者:刘鄂湖,翁泽斌,林彬燕
通讯作者单位:南京中医药大学
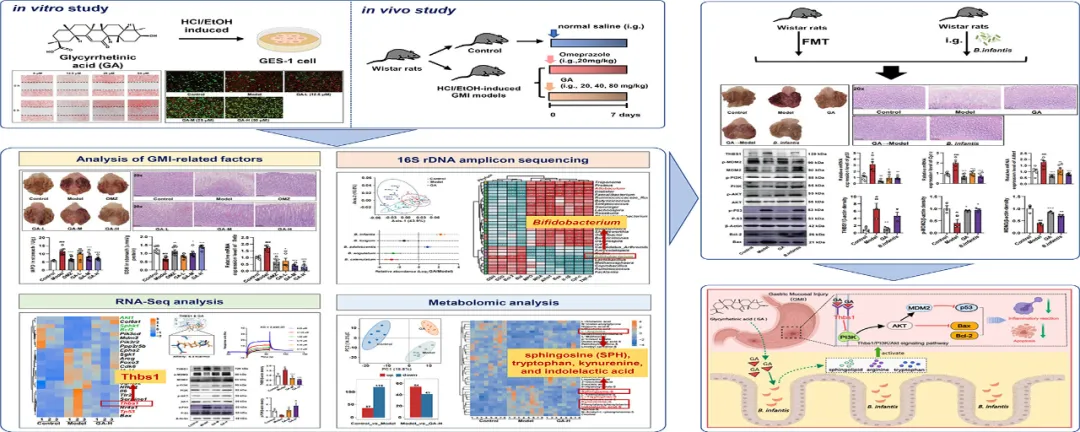
实验设计
实验结果
1. GA对HCl/EtOH诱导的GES-1细胞的影响
为了GES-1细胞开发稳定和适当的乙醇诱导损伤模型,研究者首先检查了不同乙醇浓度下3、6、12和24小时的细胞存活率(图S2A)。研究者用407.8µM乙醇孵育6小时,细胞存活率为50%(图S2B)。因此,乙醇浓度为400µM用于后续实验。然后,研究者评估了光果甘草的四种关键成分,包括甘草苷、甘草酸、GA和光甘草定对乙醇诱导的GES-1细胞损伤的保护作用。研究结果表明,GA预处理显著提高了乙醇暴露后的细胞存活率,特别是在50µM的剂量下,将细胞存活率提高到163.1±20.8%(P<0.001)(图1A-D)。在这些结果的指导下,研究者在进一步的实验中研究了GA的抗GMI作用和机制。钙黄绿素AM/PI荧光染色和膜联蛋白V/PI流式细胞术表明,与模型组相比,GA预处理(12.5、25、50µM)显著减少了GES-1细胞的凋亡(图1E-F)。细胞划痕/伤口愈合试验表明,GA在处理6小时后显著促进了GES-1细胞的迁移(图1G)。此外,12.5、25和50µM浓度的GA显著降低了HCl/EtOH诱导的GES-1细胞中Bax和p53的mRNA表达,同时显著增加了Mdm2Bcl2和Nrf2 mRNA水平(P<0.05)(图1H-I)。此外,GA显著降低了Bax与Bcl-2蛋白表达的比率,同时增加了GMI大鼠胃组织中Nrf2蛋白的水平(图1J)。
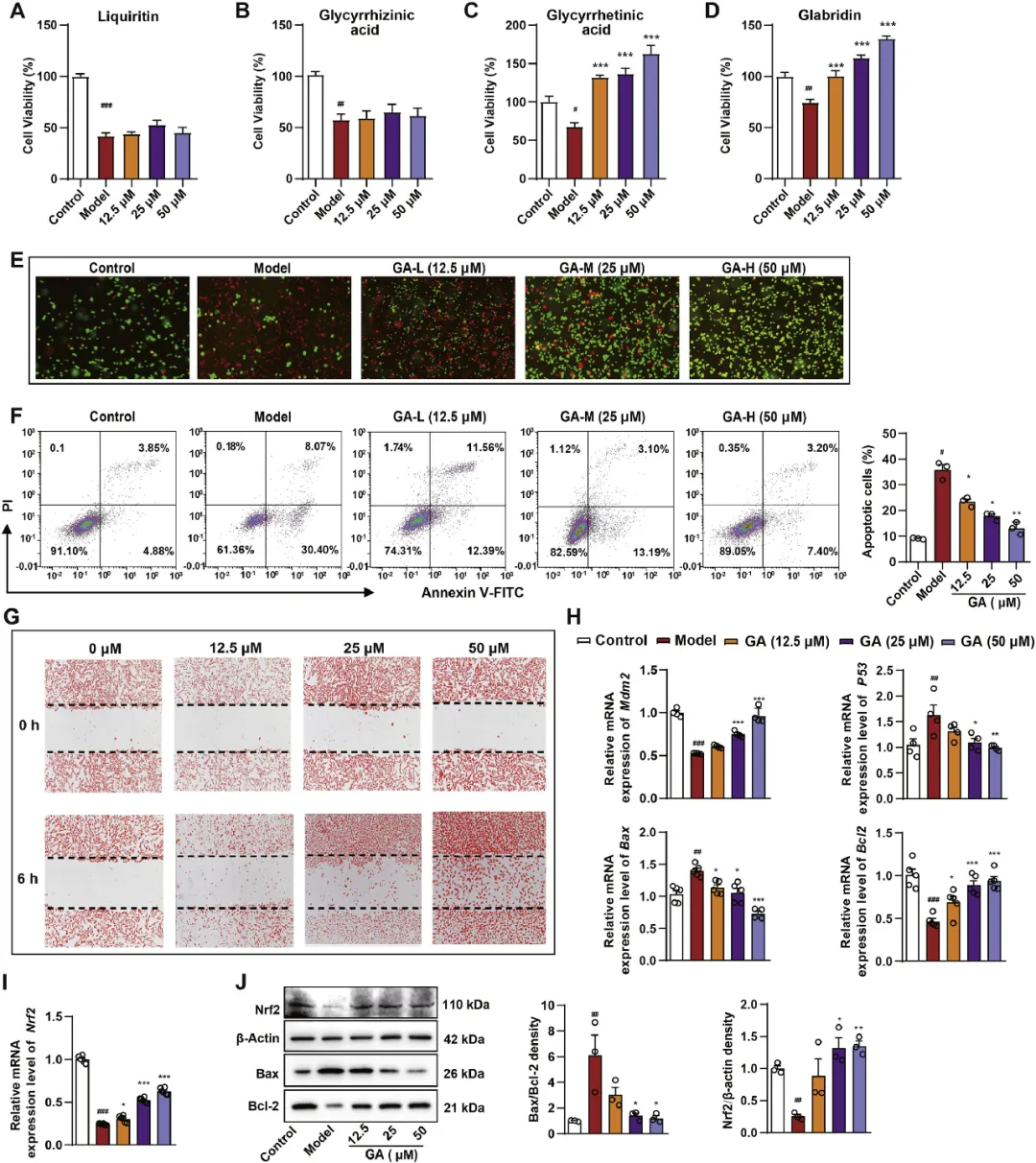
图1 GA对HCl/EtOH诱导的GES-1细胞的影响。(A-D)用不同浓度(12.5、25、50μM)的甘草苷、甘草酸、GA和光果甘定处理24小时,然后用乙醇刺激6小时后,GSE-1细胞的细胞活力。(E)用不同浓度(12.5、25、50μM)的GA(40×)处理后,活细胞(绿色)和死细胞(红色)的代表性钙黄绿素AM/PI荧光图像。(F)用不同浓度(12.5、25和50μM)的GA处理24小时,然后用乙醇刺激6小时后,用Annexin V和PI染色的细胞的代表性流式细胞术点图。(G)与对照组相比,显微照片显示了不同浓度(12.5、25、50μM)GA处理细胞前(0小时)和处理后6小时的代表性结果。(H)用不同浓度的GA处理后,GSE-1细胞中凋亡基因(Mdm2、p53、Bax和Bcl2)的相对mRNA表达,每组n=4-6。(I)用不同浓度的GA处理后,用乙醇刺激6小时,每组n=3,GSE-1细胞中Nrf2的相对蛋白表达。结果以平均值±SEM表示。(J)用不同浓度的GA处理GSE-1细胞,然后用乙醇刺激6小时后,用免疫印迹检测Nrf2、Bax和Bcl-2蛋白的表达。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
2. GA减轻了GMI大鼠的组织病理学变化
接下来,研究者评估了GA对HCl/EtOH诱导的大鼠GMI的影响。不同组别胃组织的代表性病理形态如图2A所示。与对照组相比,模型组黏膜表面出现点状或条状充血,伴有水肿和糜烂。GA-H和奥美拉唑(OMZ)组的胃组织没有充血或水肿的迹象,与对照组相似。与模型组相比,GA预处理后胃病变面积显著改善(P<0.05)(图2B)。H&E染色显示,对照组胃黏膜光滑平坦,基底上皮细胞排列紧密。相反,模型组的胃黏膜表现出明显的黏膜水肿、出血性损伤、腺体大小减小和炎性细胞浸润。与对照组相比,OMZ和GA预处理显著减轻了胃黏膜的病理损伤,导致黏膜出血减少,细胞排列更加有序。这些发现表明,GA对胃组织具有显著的保护作用(图2C)。
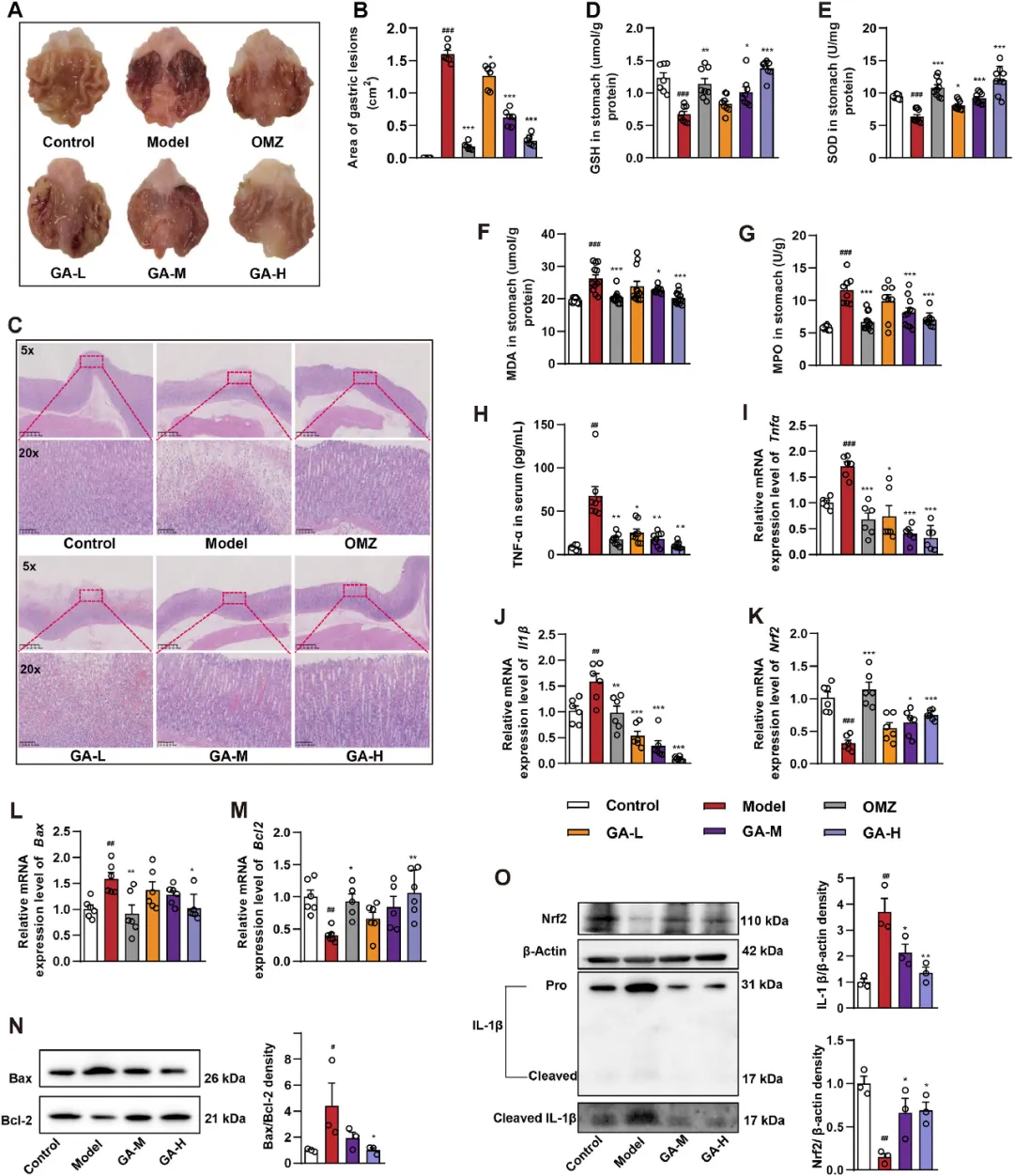
图2 GA对HCl/EtOH诱导的GMI大鼠的保护作用。将大鼠随机分为六组(n=8)。模型组(每日服用5%CMC-Na)、OMZ组(每日口服OMZ 20mg/kg/d)、GA-L/M/H组(每日口服GA 20、40和80mg/kg)。(A)不同组别大鼠的代表性胃摄影。(B)使用Image J软件对不同组的胃病变面积进行定量分析,每组n=3。(C)胃组织H&E染色的代表性显微照片(比例尺:500μm、100μm)。胃组织中氧化应激因子的水平,包括GSH(D)、SOD(E)、MDA(F),每组n=8。(G-J)血清中MPO活性、TNF-α水平和胃组织中促炎基因(TNFα和Il1β)的相对mRNA表达,每组n=6。(K-M)胃组织中Nrf2、Bax和Bcl2的相对mRNA表达,每组n=6。(N-O)免疫印迹检测GA处理的GMI大鼠胃组织中IL1β、Nrf2、Bax和Bcl-2蛋白的表达,每组N=3。结果以平均值±SEM表示。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
3. GA减轻GMI大鼠的氧化损伤和炎症反应
研究者评估了GSH、SOD和MDA水平,因为它们在氧化应激和胃溃疡中起着至关重要的作用。与模型组相比,给予GA的大鼠表现出GSH和SOD水平的显著剂量依赖性升高。GA-H和OMZ组的MDA水平显著降低,接近正常值(图2D-F)。此外,GA显著降低了MPO活性(图2G),降低了血清中TNF-α浓度,降低了胃组织中TNFα和Il1β的表达水平,OMZ组的结果相当(图2H-J)。RT-qPCR分析用于评估GA对大鼠胃黏膜组织氧化应激和凋亡相关基因的影响。在GA处理组中,Nrf2、Bid和Bcl2的表达显著增加,而Bax、Aifm1和Cyt-c的表达显著降低(图2K-M和图S3A-c)。免疫印迹分析检测GMI组织中的pro-IL1β和切割的IL-1β,结果表明,GA处理显著降低了GMI大鼠胃组织中Bax/Bcl2密度和切割的IL-1β表达,同时增加了Nrf2蛋白水平(图2N-O)。这些结果表明,GA能有效缓解HCl/EtOH诱导的炎症和氧化应激,调节凋亡相关基因的表达,保护胃黏膜。
4. GA通过调节PI3K-Akt和p53信号通路改善HCl/EtOH诱导的GMI
为了探索GA对GMI发挥保护作用的机制,研究者进行了RNA-Seq分析来评估胃基因表达谱。PCA表明,对照组、模型组和GA组的基因存在明显差异(图S4A)。研究者通过DESeq2分析,以P值<0.05和倍数变化(FC>2或FC<0.5)为标准鉴定差异表达基因(DEGs)。研究者发现,与对照组大鼠相比,GMI模型大鼠中有1468个DEGs(图S4B)发生了变化,GA大鼠中也有145个DEGs发生了变化(图S4C)。在GMI和GA处理大鼠中,95个DEGs(图S4D)共同发生了变化。其中,53个基因表现出显著趋势(图S4E),富集在与炎症、凋亡和脂质代谢相关的途径,包括PI3K-Akt、鞘氨醇和p53信号通路(图3A)。Cnetplot可视化显示了显著富集的候选途径与其相关的25个DEGs之间的重叠。节点计数超过2的基因Thbs1、Akt1、Mdm2和p53在介导GA对GMI的保护作用中起着关键的相互作用(图3B)。GA给药部分逆转了HCl/EtOH诱导的基因变化(8个上调,17个下调),使胃基因谱与对照组大鼠更接近(图3C)。实时qPCR支持RNA-seq的发现,表明GA显著降低了胃组织中Thbs1和p53的mRNA水平,同时增加了Akt1、Sphk1、Mdm2和Bcl2(图S4F和图2M)。Thbs1显示出最显著的变化,通过调节GMI发展中的PI3K、Akt、Mdm2和p53来影响炎症和凋亡。因此,GA可能通过调节Thbs1来逆转HCl/EtOH诱导的GMI大鼠效应。AutoDock结果(亲和力=-6.83±0.12 kcal/mol)表明GA通过疏水相互作用与Thbs1结合(图3D)。同时,SPR分析证实了GA对Thbs1的强结合亲和力,估计的平衡解离常数为2.46×10-7 mol/L(图3E)。免疫印迹分析显示,GA显著增加了GMI大鼠胃组织中p-MDM2、p-PI3K和p-Akt的表达,同时降低了Thbs1和p-P53的水平(图3F)。这些结果表明,GA对炎症和细胞凋亡的调节可能是由Thbs1抑制通过PI3K-Akt/p53途径触发的。
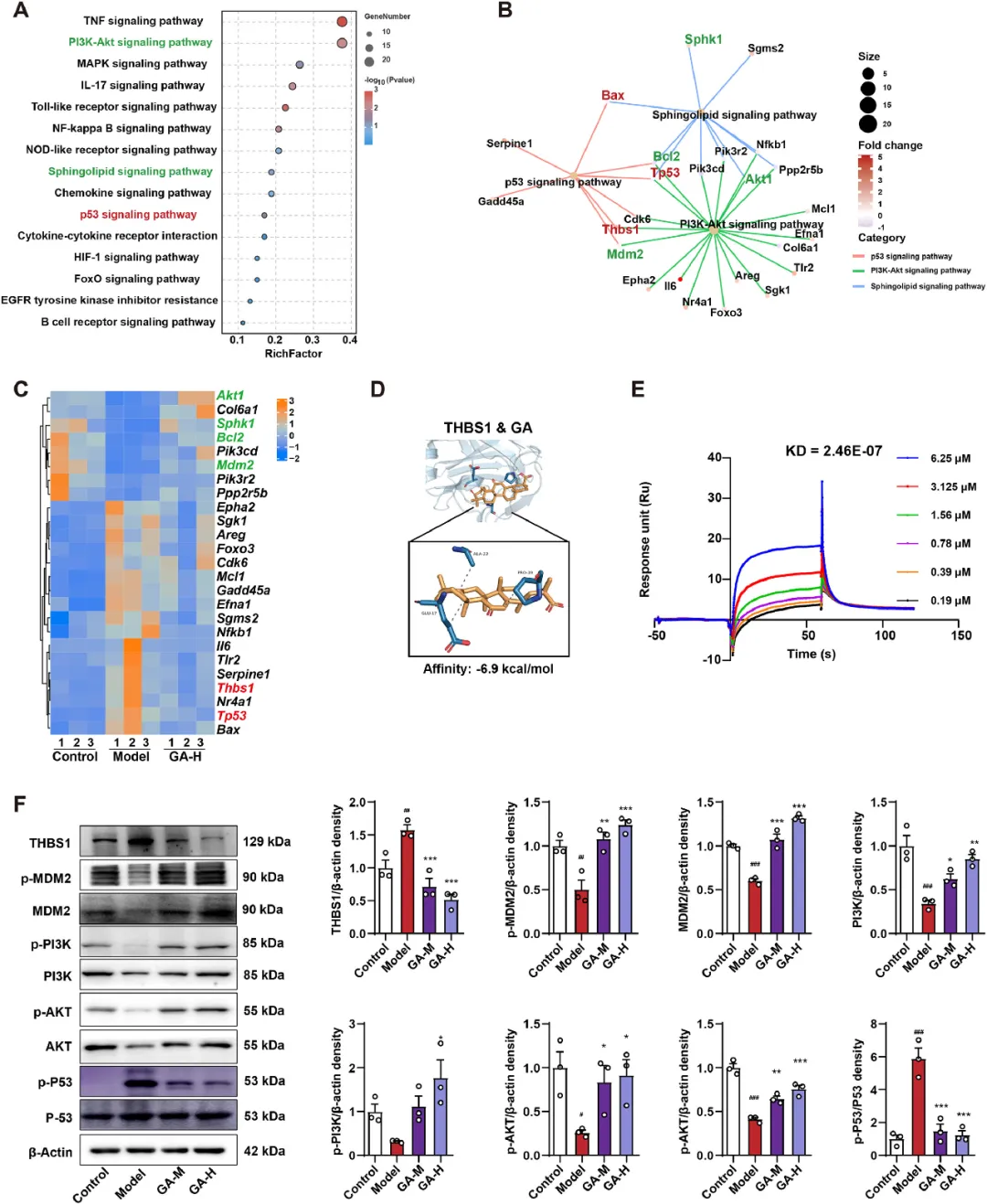
图3 GA通过调节PI3K-Akt和p53信号通路改善HCl/EtOH诱导的GMI。对照组与模型组和模型组与GA组中共同差异基因的KEGG富集分析,按显著性顺序排列(P值,TOP 15)。(B)CNEPlot可视化描绘了显著富集的候选途径与其相应的25个DEG之间的交集。(C)25个DEGs的热图,这些DEGs被HCl/EtOH改变,然后由GA调节。(D)使用AutoDock预测和分析GA分子与Thbs1的虚拟对接。(E)通过表面等离子体共振研究GA与Thbs1的结合。在注射到Thbs1生物传感器表面上时,对一系列GA浓度的频率响应进行了评估。(F)GA给药的GMI大鼠胃组织中Thbs1、MDM2、Akt、PI3K、p53和MDM2、Akt、PI3K、p53蛋白表达磷酸化的蛋白质印迹,每组n=3。结果以平均值±SEM表示。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
5. GA影响GMI大鼠肠道微生物群相关代谢产物
为了探索GA修饰的肠道微生物群对改善GMI的反应,研究者对血清和粪便样本进行了非靶向代谢组学分析。PCA显示了血清和粪便中对照组、模型组和GA组代谢物的明显分离,表明HCl/EtOH和GA处理对代谢组学特征的影响(图4A和图S5A)。如图4B-E和图S5B-E所示,OPLS-DA评分图显示了代谢谱中的明显组间分离。这些OPLS-DA模型表现出很强的分类和预测能力,截距R2Y超过0.82。粪便和血清样本中分别鉴定出215和127种代谢物,VIP值>1,P<0.05(Mann-Whitney检验)。在粪便代谢组学中(图4F-H),与正常大鼠相比,GMI大鼠的156种代谢物显示出显著差异(37种上调,119种下调),而GA显著逆转了95种代谢物(54种上调,41种下调)。同样,在血清代谢组学中(图S2F-H),研究者在GA组中观察到类似的代谢物变化。研究者通过使用在线数据库分析质谱数据,研究者确定了表S3所示的显著差异代谢物列表。GA给药部分逆转了HCl/EtOH诱导的代谢产物改变(15个上调,29个下调),导致代谢组学特征与对照组大鼠相似(图4I)。通路富集分析进一步强调,GA处理显著影响了与精氨酸生物合成、鞘脂代谢和色氨酸代谢相关的通路(图4J)。此外,研究者对各组富含精氨酸生物合成、鞘脂代谢和色氨酸代谢的差异代谢物的相对丰度进行了比较分析。在这些差异代谢物中(图4K和图S5I),GA已被证明可以降低鞘氨醇(SPH)、色氨酸、犬尿氨酸和吲哚乙酸水平,同时增加L-精氨酸和鞘氨醇1-磷酸(S1P)水平。此外,与MRS培养基中没有GA相比,GA处理的婴儿双歧杆菌显著改变了培养上清液中SPH和S1P的水平,这支持了GA处理的婴儿双歧杆菌和鞘氨醇代谢物之间的直接联系(图4L)。
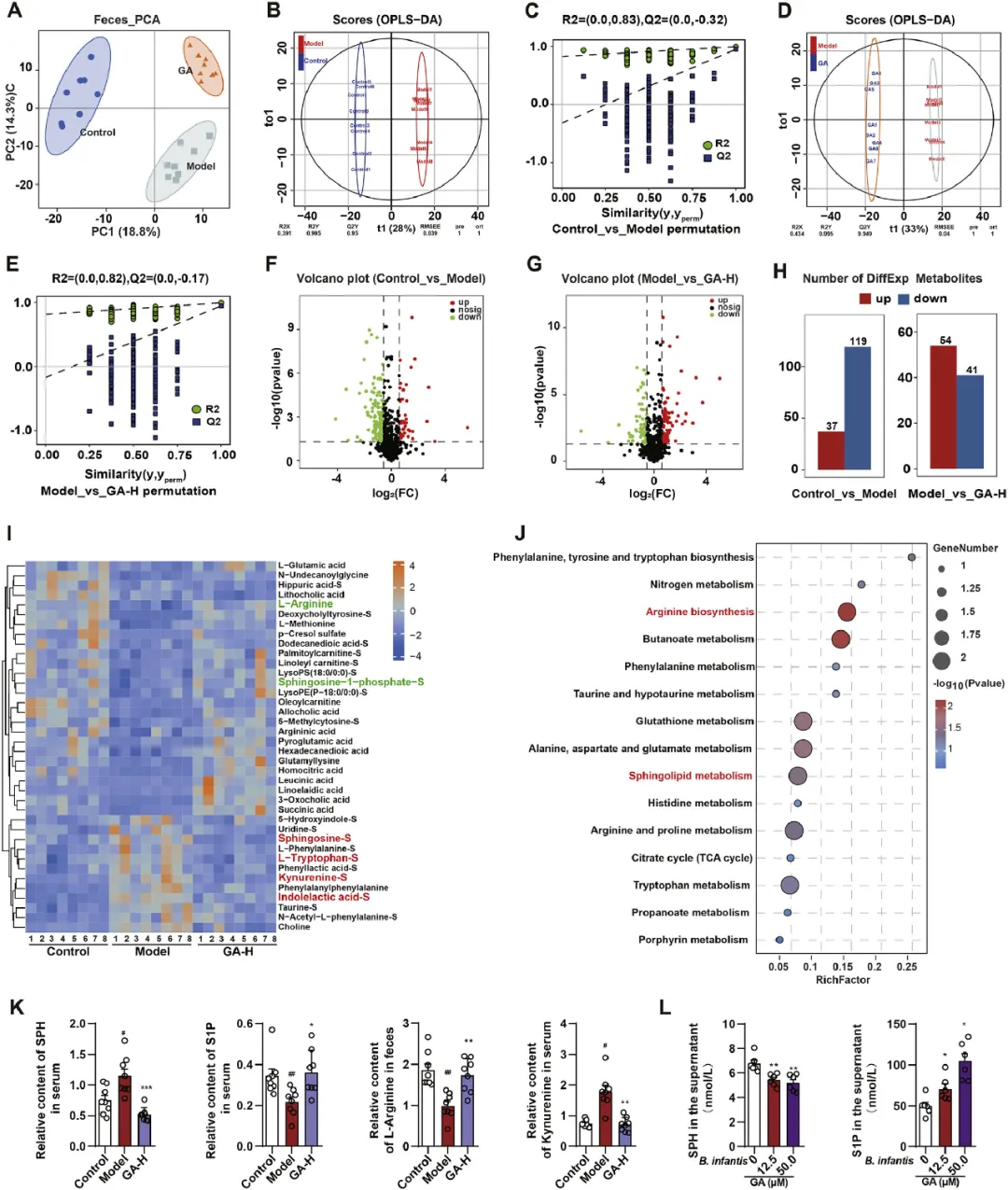
图4 GA影响与GMI大鼠肠道微生物群相关的代谢物。对对照组、模型组和GA-H(80mg/kg)组的血清和粪便样本进行代谢组学分析,这些样本是在第7天使用UHPLC-QTOF/MS收集的。来自(A)主成分分析(PCA)评分图的血清样本的PCA评分图,用于区分粪便代谢组与对照组、模型组和GA组。(B-E)对照与模型(粪便)、模型与GA(粪便)的OPLS-DA评分图以及相应的置换检验。通过OPLS-DA和Mann-Whitney检验,对照组与模型组、模型组与GA组大鼠粪便代谢组学的火山图显示了VIP值>1和P<0.05的代谢物异常值。(H)对照组与模型组、模型组与GA组的差异代谢物数量。(I)由HCl/EtOH改变然后由GA调节的差异代谢物的热图。(J)差异代谢物的前15个富集KEGG术语通路,通路分析由MetaboAnalyst 5进行。(K)各组间富含精氨酸生物合成、鞘脂代谢和色氨酸代谢的差异代谢物(SPH、S1P、L-Arg、犬尿氨酸)的相对丰度,每组n=8。(L)GA处理婴儿双歧杆菌培养上清液中SPH和S1P的ELISA分析,每组n=6。结果以平均值±SEM表示。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
6.GA改变了GMI大鼠肠道微生物群的组成和功能
考虑到肠道微生物群在鞘脂和色氨酸代谢中的作用,研究者通过对对照组、模型组和GA处理组的粪便样本进行16S rRNA基因测序,研究了GA对肠道微生物群的影响。与模型组相比,GA组的Chao1和Shannon指数显著降低,表明GA处理大鼠的生物多样性和丰富度降低(图5A)。PCA清楚地区分了三组之间的肠道微生物群特征,突出了HCl/EtOH和GA引起的显著变化(图5B)。在属水平上,模型组具有较高的Allobaculum、Dubosiella和Muribaculaceae丰度,但乳杆菌、双歧杆菌和布劳特氏菌属丰度较低。GA处理改变了肠道微生物组成,减少了Allobaculum、Dubosiella和Muribaculaceae,同时增加了乳杆菌属、双歧杆菌属和布劳特氏菌属(图5C)。LDA评分(log10)>3表明优势菌株在每组中的相对表达增加。模型组的微生物群落与对照组存在显著差异,Allobaculum、Oscillospira、Proteus和Phascolarctobacterium的水平升高。GA显著影响了肠道微生物群中的特定细菌种类。与模型组相比,GA组显示双歧杆菌和瘤胃球菌显著富集(图5D)。这些发现表明,GA调节GMI大鼠的肠道微生物群组成。Spearman的相关性分析显示,MPO、MDA、Aifm1、Bax、Tnfα、Cyt-c基因与Allobaculum和Dialister呈正相关。相反,Allobaculum和Dialister与Bcl2和Bid基因呈负相关。双歧杆菌与MDA、Aifm1、Bax、Il1β、Tnfα和Cyt-c基因呈负相关,与GSH、SOD、Bcl2和Bid基因呈正相关(图5E),表明双歧杆菌的富集和Allobaculum丰度的减少可能在GA对HCl/EtOH诱导的GMI的保护作用中发挥作用(图5F)。qPCR分析显示,GA增加了B.infantis、B.longum和B.adolescentis的水平,同时降低了B.angulatum和B.catenulatum的水平,其中B.infanti的富集程度最高(图5G)。这些发现表明,GA对GMI的保护作用可能与婴儿双歧杆菌有关。
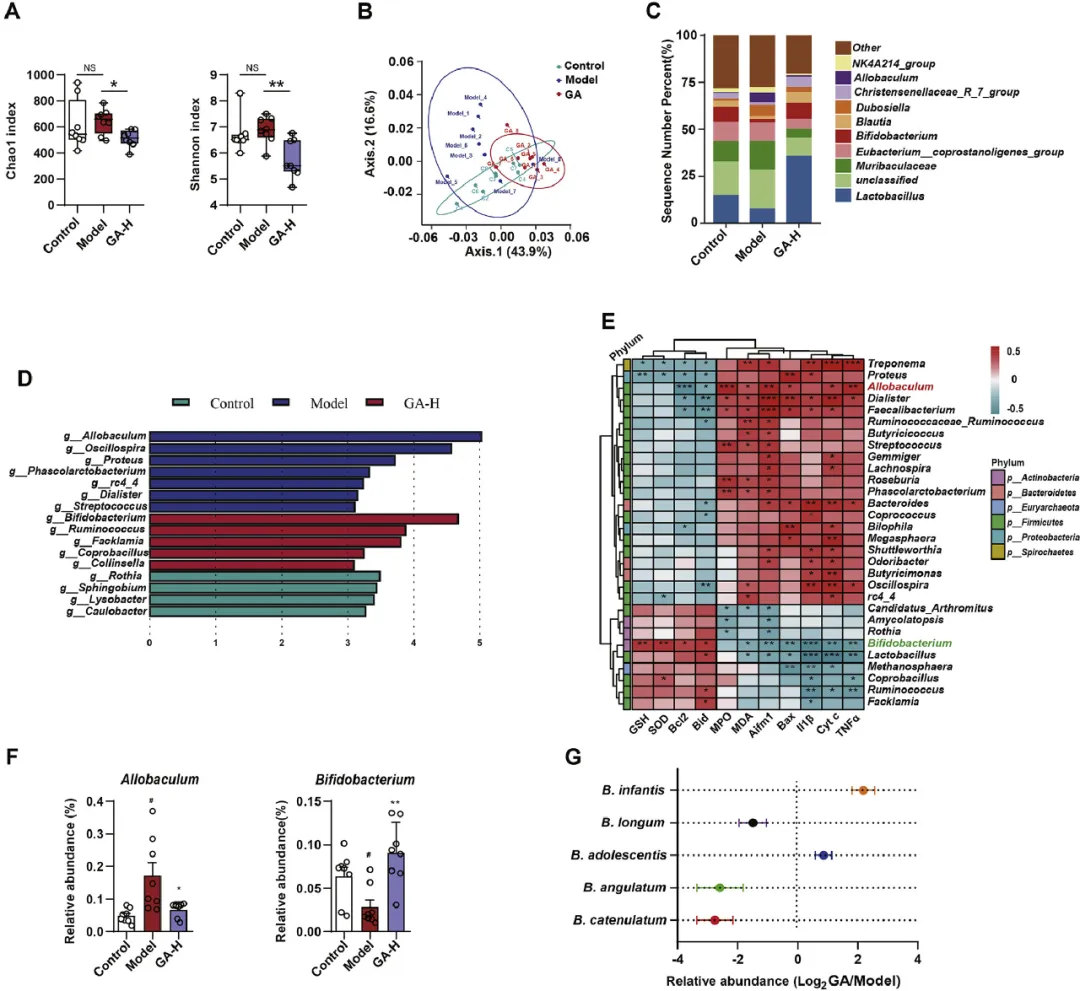
图5 GA改善了HCl/EtOH诱导的GMI大鼠的肠道微生物群失调。通过16S rRNA测序分析第7天收集的对照组、模型组和GA-H(80 mg/kg)组的粪便中的肠道微生物群,每组n=8。(A)不同组肠道微生物群的Chao多样性和Shannon多样性,(B)加权UniFrac-PCoA分析。(C)属水平的细菌分类分析。(D)通过LEfSe(log10-LDA>3.0)确定鉴别分类群。(E)Kruskal-Wallis H检验显示,30个属之间的显著差异热图(P<0.05),这些差异被HCl/EtOH改变,然后被GA调节;代谢产物和GMI相关症状的Pearson相关热图分析具有统计学意义,P值<0.05。(F)GA处理后HCl/EtOH诱导的GMI大鼠Allobaculum和双歧杆菌的相对丰度。(G)指定组粪便中双歧杆菌物种的相对DNA表达(以Log2 GA/模型表示)。结果以平均值±SEM表示。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
7. GA可以通过丰富婴儿双歧杆菌来缓解GMI
为了进一步验证GA通过富集婴儿双歧杆菌来增强GMI的影响,研究者将GA-H组的粪便样本移植到FMT组(伪无菌大鼠),以评估GA是否通过肠道微生物群调节来改善HCl/EtOH诱导的大鼠GMI。此外,GMI大鼠连续七天每天服用婴儿双歧杆菌(1×1010CFUs/kg)。胃组织的形态学分析和H&E染色显示,FMT和婴儿双歧杆菌均显著减轻了GMI,表现为炎性细胞浸润、黏膜水肿和上皮细胞分离减少(图6A-C)。此外,与模型组相比,GA、FMT和婴儿双歧杆菌治疗组的大鼠GSH、SOD、Nrf2、Mdm2和Bcl2水平显著升高,MDA、MPO、TNF-α、Il1β、p53、Cyt-c、Aifm1和Bax水平显著降低(图6D-N和图S6A-B)。这些发现表明,GA对GMI的有益作用可以通过FMT有效重现。随后的分析表明,婴儿双歧杆菌的给药显著逆转了GMI大鼠炎症和凋亡相关蛋白的表达,如IL-1β、Thbs1、Nrf2、p-MDM2、p-PI3K、p-AKT、p-P53、Bax和Bcl-2(图6O-p),强调GA在GMI大鼠中的抗炎作用是由婴儿双歧杆菌的富集介导的。这些结果表明,肠道微生物群在介导GA的保护作用中起着关键作用,强调了大鼠肠道微生物群的这些变化对GA的抗GMI特性有显著影响。
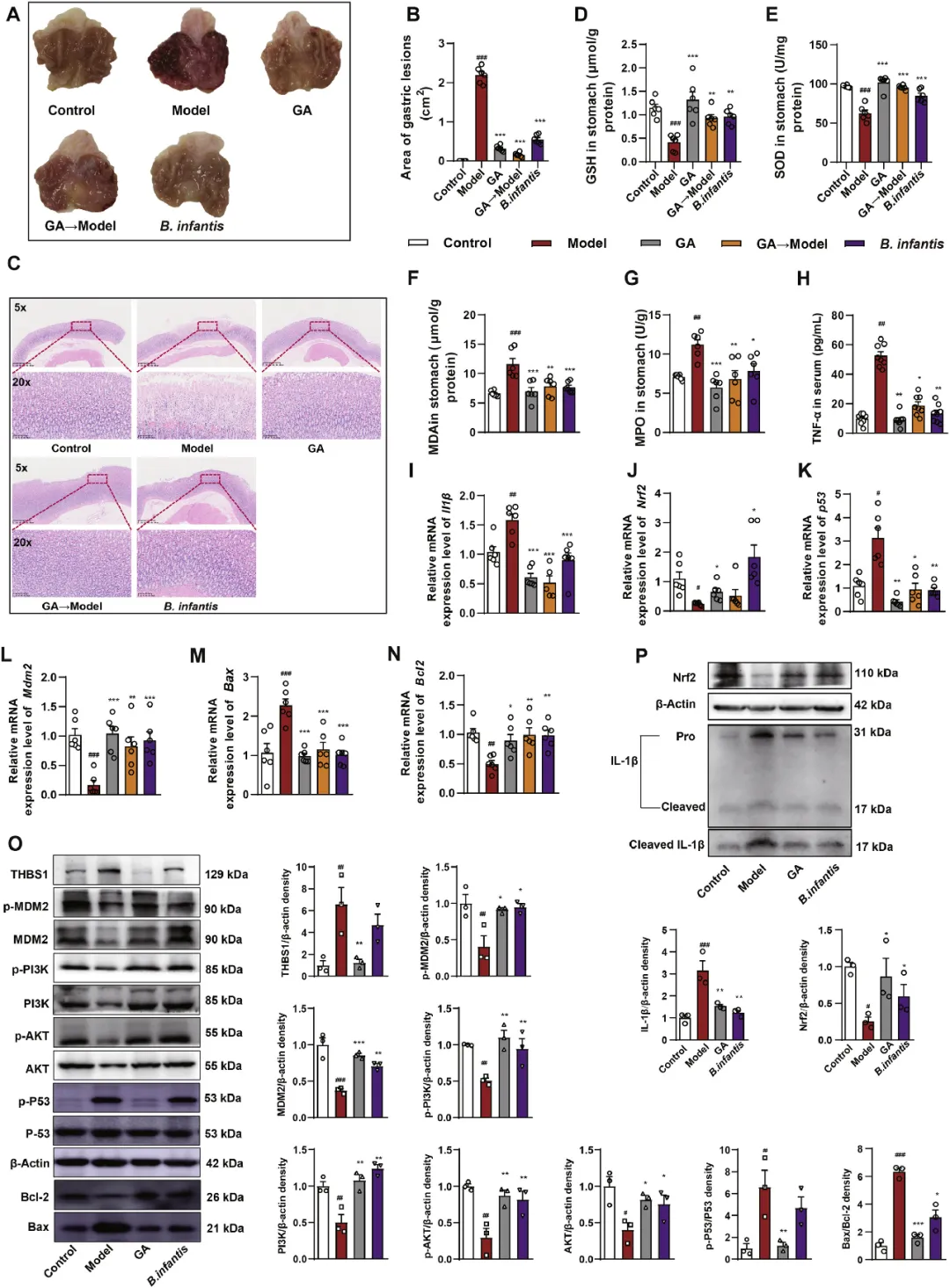
图6 GA可以通过丰富婴儿双歧杆菌来缓解GMI。将大鼠随机分为5组(n=8)。模型组(每日口服5%CMC-Na)、GA组(每日服用GA 80mg/kg)、GA模型组(每天口服GA组的微生物群)、婴儿双歧杆菌(每日口服1×1010 CFUs/kg婴儿双歧杆菌)。(A)不同组别大鼠的代表性胃摄影。(B)使用Image J软件对不同组的胃病变面积进行定量分析,每组n=3。(C)胃组织H&E染色的代表性显微照片(比例尺:500μm、100μm)。胃组织中氧化应激因子的水平,包括GSH(D)、SOD(E)、MDA(F),每组n=6。(G-I)血清中MPO活性、TNF-α水平和胃组织中促炎基因(Il1β)的相对mRNA表达,每组n=6。(J-N)胃组织中基因(Nrf2、p53、Mdm2、Bax和Bcl2)的相对mRNA表达,每组N=6。(P-O)用免疫印迹检测GA或B. infantis治疗的GMI大鼠胃组织中IL-1β、Nrf2、Thbs1、MDM2、Akt、PI3K、p53、Bax、Bcl-2和MDM2、Akt、PI3K、p53蛋白表达的磷酸化,每组n=3。结果以平均值±SEM表示。P值通过Dunnett事后检验的单因素方差分析确定。与对照组相比:#P<0.05,##P<0.01,###P<0.001;与模型组相比:*P<0.05,**P<0.01,***P<0.001。
讨论
HCl/EtOH诱导的GMI模型是一种广泛认可的临床前工具,用于研究急性黏膜损伤的机制和评估潜在的保护剂。通过模拟胃酸分泌过多和酒精滥用,HCl和乙醇通过黏液糖蛋白变性、ROS升高和促进中性粒细胞浸润,协同破坏胃黏膜屏障。这些病理过程与人类GMI的病因非常相似,包括酒精诱导的胃炎、非甾体抗炎药相关的损伤和应激性溃疡。HCl/EtOH诱导的胃损伤的病理生理学包括炎症、凋亡细胞死亡和胃中的氧化应激。HCl/EtOH显著升高MPO、Tnfα和Il1β的水平,通过促进中性粒细胞浸润和激活氧化应激、有毒代谢物和溶酶体酶加剧组织损伤,导致局部组织损伤。此外,它会导致过量自由基和脂质过氧化物的积累,对蛋白质、脂质和DNA产生不利影响,最终导致细胞死亡。评估胃组织氧化应激的关键标志物包括MDA、GSH、SOD和Nrf2。HCl/EtOH处理显著增加了MDA水平,同时降低了GSH和SOD活性。在这种情况下,HCl/EtOH增强了黏膜中的促凋亡事件,例如Bax、Aifm1和Cyt-c的过表达mRNA水平,同时下调了抗凋亡Bcl-2和Bid。Nrf2-Keap1通路调节氧化应激和细胞氧化还原平衡,在胃黏膜保护中起着重要作用。在活性氧(ROS)积累后,活化的Nrf2与抗氧化反应元件相互作用,上调抗氧化和解毒基因的表达,包括HO-1、NQO1和GSH。这增强了细胞消除ROS和修复氧化损伤的能力。本研究结果表明,GA显著提高了HCl/EtOH诱导的GES-1细胞和GMI大鼠中Nrf2的表达,表明GA有效地减轻了HCl/EtOH暴露引起的氧化应激。综上所述,本研究结果表明,GA下调炎症因子和凋亡相关基因,同时降低氧化应激水平。
肿瘤抑制蛋白p53通过上调Bax和抑制Bcl-2表达来抑制细胞生长并诱导凋亡。磷酸化和去磷酸化对于调节这些过程至关重要。磷酸化稳定Mdm2,使其能够结合p53并促进其泛素化和降解。此外,磷酸化在体内和体外都会减少p53与其负调控因子Mdm2之间的相互作用。在本研究中,研究者证明GA和婴儿双歧杆菌在抑制p53功能的同时增加了Mdm2磷酸化。这一过程下调Bax,上调Bcl-2,促进细胞增殖,同时减少凋亡。
微生物群组成和多样性的改变与胃肠道疾病密切相关。通过粪便微生物群移植试验,研究者发现将GA处理的大鼠的肠道微生物群转移到微生物群耗尽的大鼠身上可以改善GMI的各个方面,如减少氧化应激、降低凋亡基因的表达和降低炎症因子水平。这些结果表明,GA通过调节肠道微生物群来增强GMI。为了鉴定受GA影响的特定肠道细菌,研究者收集了大鼠的粪便样本,并进行了16S rRNA测序以评估微生物群组成。在属水平上,与模型组相比,GA通过显著增加双歧杆菌等有益菌来抵消HCl/EtOH诱导的破坏。其中,婴儿双歧杆菌被认为通过调节免疫反应和治疗炎症性肠病来增强胃肠道健康。研究者的数据表明,与对照组相比,婴儿双歧杆菌显著改善了大鼠的胃黏膜损伤。在化疗诱导的肠黏膜炎中,婴儿双歧杆菌通过调节细胞因子表达和分化因子来降低Th1细胞反应。它还能有效抑制NF-κB信号通路的激活,从而减轻炎症。此外,其生物活性肽在体内显示出活性,恢复肠道通透性,缓解结肠炎。这些发现强调,GA限制了病原体的增殖,增强了大鼠肠道中的有益菌,减少了炎症,纠正了微生物群的不平衡。此外,长双歧杆菌CECT7894的应用改变了与鞘脂代谢相关的基因表达、代谢物水平和肠道微生物群的相对丰度,从而调节了肠道稳态和整体宿主健康。总的来说,通过促进婴儿双歧杆菌的定植,GA有可能调节肠道微生物群的组成,从而通过色氨酸和鞘脂代谢影响S1P、精氨酸和色氨酸代谢物(犬尿氨酸和吲哚乳酸)的水平,这有助于抑制炎症和改善胃肠道完整性。上述结果表明,GMI的治疗策略很有前景。
鉴于肠道细菌通过各种代谢物调节身体功能的重要作用,研究者进行了彻底的血清和粪便代谢组学分析。在质膜上,鞘脂转化为鞘氨醇(SPH),然后被鞘氨醇激酶(SPHK)转化为S1P。S1P也是一种溶血磷脂,在许多信号转导途径中起着至关重要的作用,影响着各种细胞过程,如血管内皮细胞功能、炎症、凝血、血栓形成、细胞生长和凋亡。此外,它通过与S1PR1受体相互作用,激活PI3K/Akt通路,增强内皮祖细胞的增殖和抗凋亡作用。因此,研究者提出GA可以通过富集BI细菌来驱动SPH-SIP的代谢重建并调节PI3K-AKT通路,从而缓解乙醇诱导的胃溃疡。此外,L-精氨酸通过相同的PI3K-Akt通路在凋亡调节中起着关键作用,并通过抑制NF-κB和MAPK通路的激活来减轻炎症和肠道损伤。同样,色氨酸是许多微生物和宿主代谢物生物合成所必需的,在胃肠道中进行代谢。本研究结果证实,GA可以缓解色氨酸及其衍生物(犬尿氨酸和吲哚乙酸)水平的升高,从而调节HCl/EtOH诱导的GMI大鼠的炎症反应。这些见解表明,鞘脂和色氨酸代谢是GA提供的GMI保护机制不可或缺的一部分。
凝血酶敏感蛋白-1(Thbs1)最初在血小板中被鉴定为一种粘附性糖蛋白,具有多种生物学功能,包括抗血管生成、凋亡调节、TGF-β激活和免疫调节。研究者发现GA可以抵消HCl/EtOH诱导的GMI大鼠胃组织中Thbs1的过表达。然而,GA如何影响这种保护仍有待澄清,特别是它是否靶向Thbs1。研究者的探索通过SPR和分子对接研究证实了GA与Thbs1的强结合亲和力。THBS1的凝血酶敏感蛋白重复序列通过与CD36和CD47结合并调节PI3K/Akt通路来抑制内皮细胞迁移并促进凋亡。因此,研究者假设Thbs1的下调对HCl/EtOH处理的GMI大鼠中GA的保护机制至关重要,这是通过调节Thbs1/PI3K-Akt/p53信号通路介导的。总之,本研究证实了GA对胃黏膜的保护作用,通过PI3K-Akt/MDM2/p53信号通路证明了其抗凋亡能力。有效减轻炎症和氧化应激,促进胃黏膜屏障的修复。此外,FMT实验强调了肠道微生物群在调节GA疗效中的关键作用。通过16S rRNA测序和代谢组学分析,研究者发现GA缓解了HCl/EtOH诱导的肠道失调,并解决了鞘氨醇、色氨酸和精氨酸代谢的不平衡问题。通过增强肠道微生物群中的婴儿双歧杆菌,GA有效地纠正了与黏膜损伤相关的代谢紊乱。此外,其机制可能涉及抑制Thbs1/PI3K-Akt/p53信号通路(图7)。未来的研究对于进一步阐明GA和Thbs1之间的因果关系至关重要。具体而言,研究应调查GA及其相关代谢产物SIP是否可以直接调节Thbs1的表达,并影响PI3K/AKT信号通路介导的保护机制。
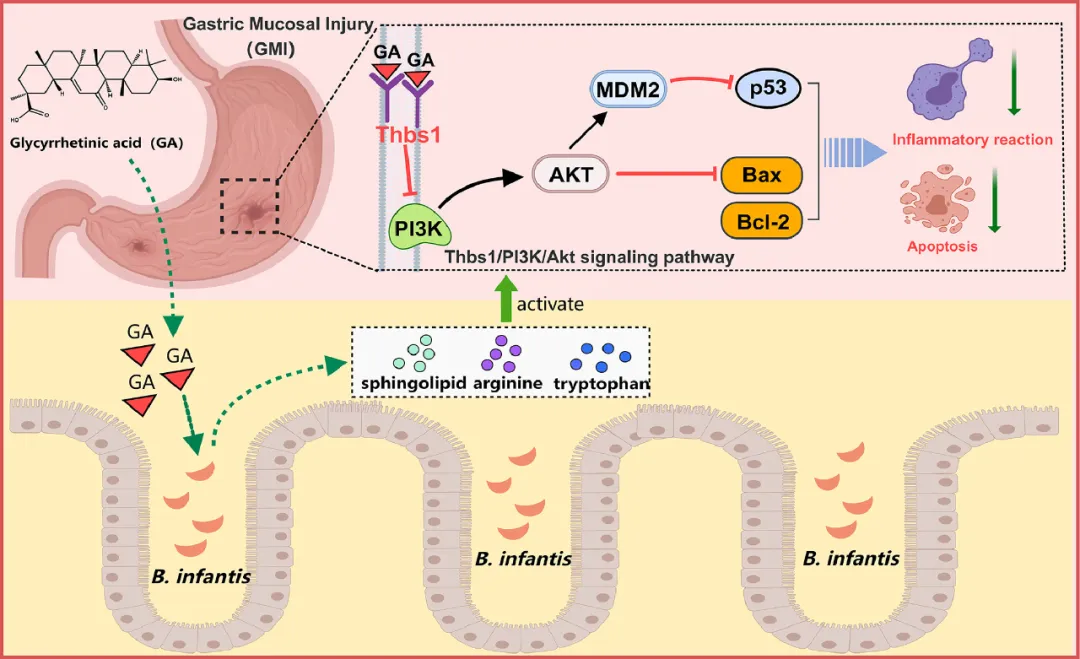
图7 甘草次酸通过Thbs1/PI3K-Akt/p53途径调节肠道微生物群及其代谢产物来改善胃黏膜损伤。
结论
总之,本研究表明,GA给药通过改善HCl/EtOH诱导的GMI大鼠的肠道生态失调和促进婴儿双歧杆菌定植来调节肠道微生物群的组成。它调节精氨酸和色氨酸水平,影响色氨酸代谢。此外,GA已显示出作为调节p53和PI3K-Akt信号通路的治疗剂的潜力,这有助于减轻与肠道微生物群失衡相关的炎症反应。
科研 | 长春中医大:野山参中低极性人参皂苷通过肠道菌群驱动色氨酸代谢激活AhR/MAPK通路提高免疫力(国人佳作)
科研 | 天中医:泽泻多糖通过调节肝脏代谢和肠道菌群改善对乙酰氨基酚引起的急性肝损伤(国人佳作)
如果需要原文pdf,请扫描文末二维码



获取此文献原文PDF、申请加入学术群,联系您所添加的任一微科盟组学老师即可,如未添加过微科盟组学老师,请联系微生态老师10,无需重复添加。
请关注下方名片,了解更多代谢组学知识
随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 国际名校科研合作!博士后/专职科研岗,南京大学科研团队招聘,2026年8月8日截止报名
- 不靠区划扩容做支撑,南京凭科教产业硬实力,稳居长三角特大城市前列.
- 南京今天·2026年5月8日
- 【5月21日-南京】第219届拨针、刺骨等微创技术治疗骨伤、关节病培训班
- 南京公益性岗位招聘 | 招17人!南京白下人力资源开发服务有限公司招聘劳务派遣人员公告(十)
- 叶帅来南京,许世友拄拐上街迎候,两小时问了秘书5次:到了没有
- 总分408!南京大学新闻与传播考研经验|高分锦囊
- 南京市2026届高三英语二模作文可以这么写
- 贤·园 | 南京市七贤幼儿园2026年秋季招生公告
- 南京警察学院438警务硕士专业综合2026年考研真题及答案(回忆版,只有公基部分)
