维持氧化还原稳态对于胶质母细胞瘤(GBM)等侵袭性癌症的存活、进展及治疗耐药性至关重要。GBM是最常见且最致命的原发性脑肿瘤之一,具有快速增殖、侵袭性强及对常规治疗耐药的特点。胶质瘤细胞的一个显著特征是它们在氧化应激下能够维持氧化还原稳态,这主要依赖于内源性抗氧化系统的上调,尤其是谷胱甘肽(GSH)。GSH可清除活性氧(ROS)、维持增殖信号传导,并增强治疗耐药性。这一氧化还原调控系统与膜受体信号网络紧密耦联,其中G蛋白偶联受体(GPCRs)发挥着核心作用。GPCRs是最大的膜受体家族,对于将细胞外信号转导为细胞内反应至关重要。
β-肾上腺素能受体(β-ARs)是GPCRs的一个重要亚类,在心血管功能和代谢稳态中发挥关键调控作用。在三种β-AR亚型(β1、β2和β3)中,β3-AR已成为氧化还原生物学的重要调节因子。该受体广泛分布于多种组织,包括脑组织,并因其与多种生理过程(尤其是炎症和氧化应激)的关联而日益受到关注。研究表明,激活β3-AR可通过上调NADPH氧化酶活性并促进抗氧化酶的表达来调节细胞氧化还原稳态,从而与细胞氧化应激反应建立功能性联系。值得注意的是,β3-AR在高级别胶质瘤中过表达,提示其参与肿瘤对氧化应激的适应及恶性进展。这些观察结果提出一个令人信服的假说:胶质瘤细胞可能利用β3-AR在氧化还原调控中的已知功能来维持自身存活,提示靶向β3-AR以破坏氧化还原平衡具有治疗潜力。然而,β3-AR信号传导与胶质瘤氧化还原稳态之间的确切机制联系仍不明确。一个主要障碍在于缺乏可穿透血脑屏障(BBB)的分子工具,无法在天然完整的脑环境中同时实时原位可视化β3-AR的分布并监测周围的氧化还原状态(尤其是GSH水平)。传统成像方法不足以探测大脑复杂肿瘤微环境中受体的空间分布和动态氧化还原波动。
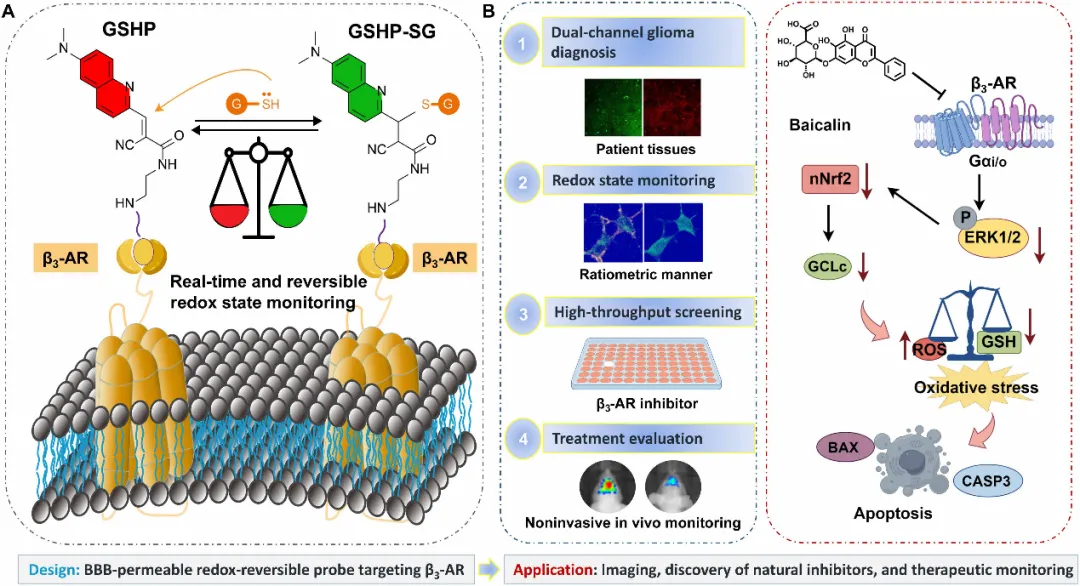
在此,作者报道了GSHP——一种经合理设计的、具有血脑屏障通透性的双功能荧光探针,能够同时实现β3-AR分布的成像和局部谷胱甘肽(GSH)波动的实时监测(图1)。GSHP的设计将源于卡拉洛尔“弹头”结构的β3-AR靶向基团整合到可穿透血脑屏障的喹啉荧光骨架中,并引入一个分子转子单元,该单元在与受体结合时产生增强的荧光信号,从而提供β3-AR分布的空间读数。同时,其含氰基的不饱和烯基部分与局部GSH发生可逆的迈克尔加成反应,产生显著蓝移的荧光发射。这种双通道荧光响应使作者能够特异性地评估β3-AR富集的胶质瘤微环境内的GSH波动,从而为局部的氧化还原动态提供独特的见解。此外,利用GSHP进行高通量筛选,作者发现并验证了天然化合物黄芩苷是一种β3-AR天然抑制剂,它通过Gαi/o-ERK-Nrf2-GCLc信号通路,在U251胶质瘤细胞中消耗GSH、诱导氧化应激并触发细胞凋亡。在原位胶质瘤模型中,借助GSHP的血脑屏障穿透能力,作者使用双通道GSHP成像无创地绘制了β3-AR表达模式和局部GSH梯度,实现了肿瘤的高对比度描绘,并为临床前模型中胶质瘤治疗效果的无创监测与评估提供了一种工具。该平台有望为阐明β3-AR在胶质瘤生物学和氧化还原调控中的作用开辟新途径,并促进靶向脑肿瘤氧化还原敏感通路的天然调节剂或相关治疗分子的发现。
图1. GSHP——一种具有血脑屏障通透性、靶向β3-AR的氧化还原可逆探针的设计与应用示意图。(A)GSHP是一种双响应荧光探针,能够同时靶向并成像β3-AR,同时可逆地监测受体局部微环境中的GSH稳态变化。(B)该多功能探针可用于双通道胶质瘤诊断、氧化还原状态监测、高通量药物筛选以及胶质瘤治疗效果评估。深入的机制研究表明,通过GSHP筛选鉴定出的黄芩苷是β3-AR的一种天然抑制剂。通过抑制β3-AR及其下游的Gαi/o-ERK-Nrf2-GCLc信号通路,黄芩苷可耗竭U251 GBM细胞中的GSH,从而诱导氧化应激介导的细胞凋亡。
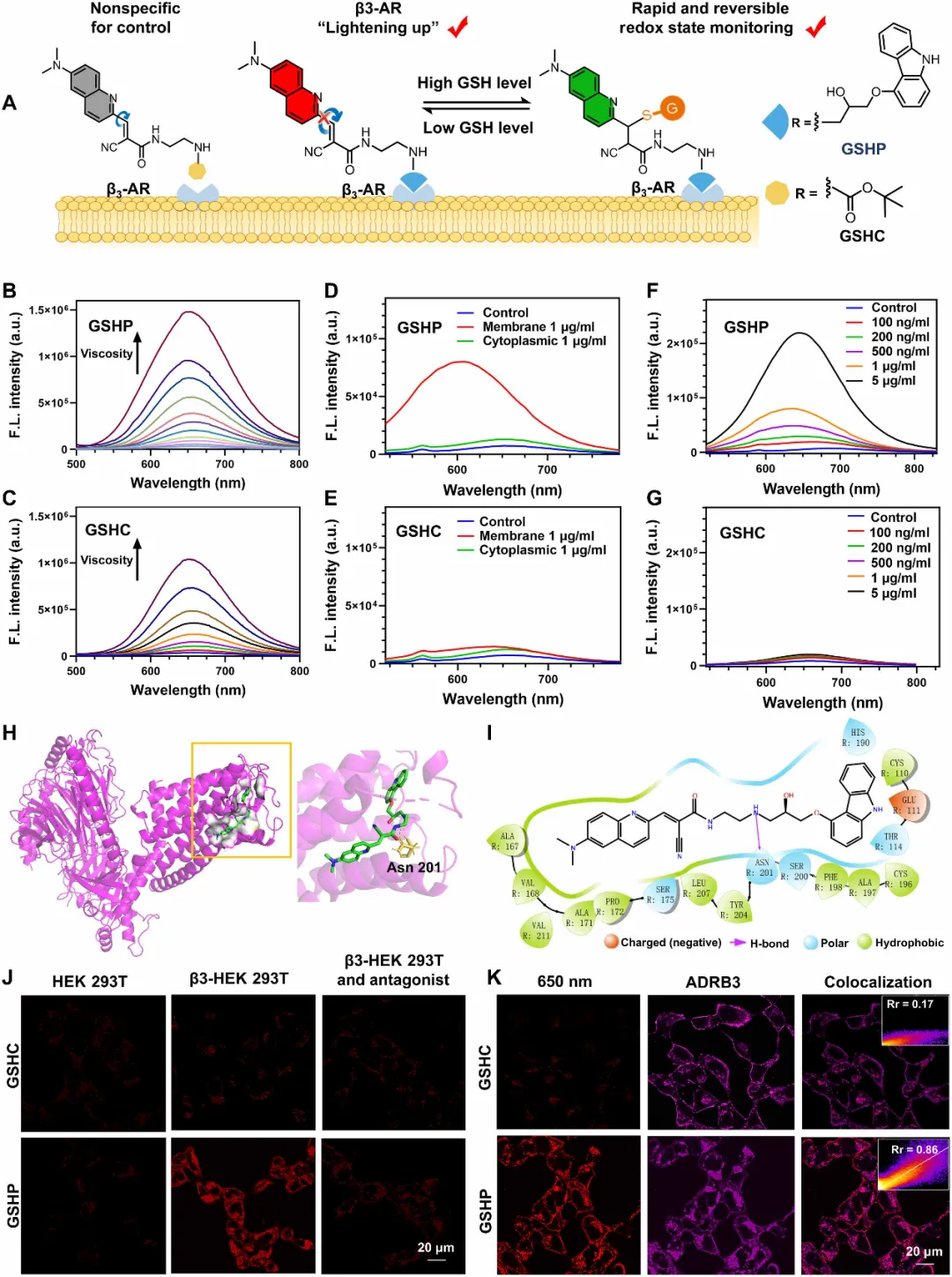
图2. 用于β3-AR成像和氧化还原监测的双响应荧光探针GSHP的设计。(A)GSHP将β3-AR靶向功能与GSH响应信号相结合,能够同时实现受体分布和局部氧化还原动态的可视化。GSHC(缺少靶向基团)作为非特异性对照探针。(B和C)GSHP(B)和GSHC(C)(各10 μM)在不同粘度的PBS/甘油混合液中的荧光光谱。(D和E)在从过表达β3-AR的HEK293细胞提取的胞质蛋白或膜蛋白(各1 μg/ml)存在下,GSHP(D)和GSHC(E)(各1 μM)的荧光光谱(λex = 470 nm,λem = 655 nm,狭缝=5/5 nm,pH 7.4,37°C)。(F和G)随着膜蛋白浓度增加(0至5 μg/ml),GSHP(F)和GSHC(G)(各1 μM)的荧光光谱。(H)GSHP与β3-AR(PDB:7XJH)的分子对接,显示整体结合姿态及与残基Asn201的相互作用。(I)GSHP在β3-AR结合口袋内的二维相互作用图。(J)将HEK293T细胞和转染β3-AR的HEK293T细胞与GSHP或GSHC(100 nM)共同孵育,在存在或不存在β3-AR拮抗剂卡拉洛尔的情况下的荧光成像(λex = 488 nm,λem = 620–680 nm)。(K)将转染β3-AR的HEK293T细胞与GSHP或GSHC(100 nM)共同孵育后的免疫荧光成像。β3-AR通道(ADRB3)以伪品红色显示(λex = 561 nm,λem = 580–620 nm)。a.u.,任意单位。
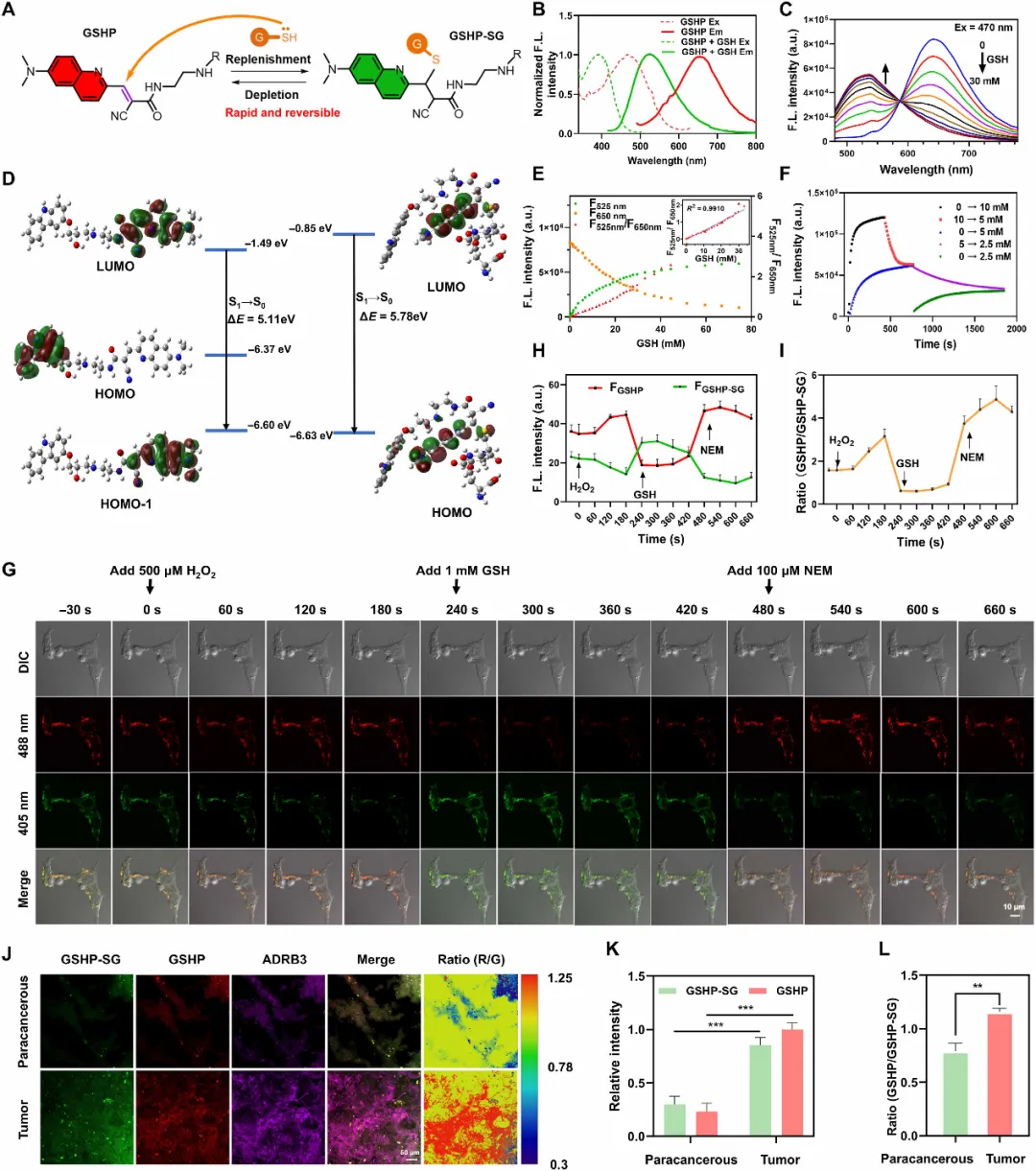
图3. GSHP的可逆荧光响应及其在胶质瘤细胞和组织中的成像应用。(A)GSHP与GSH之间可逆反应的示意图。(B-F)GSHP(10 μM)对GSH的比率荧光响应的体外表征。(B)加入GSH(10 mM)之前(红色)和之后(绿色)的归一化激发/发射光谱。(C)随GSH浓度增加(0 至 30 mM)的荧光滴定光谱,显示比率变化。(D)通过DFT计算得到的GSHP及其加成物的前线分子轨道。(E)荧光比值(F525/F650)随GSH浓度增加而升高,在0–30 mM范围内呈良好线性(插图,R² = 0.991)。(F)动力学分析证实反应的可逆性。(G-I)负载GSHP的U251细胞的活细胞成像,显示对依次加入H₂O₂(500 μM)、GSH(1 mM)和NEM(100 μM)的动态比率响应(G);各通道荧光强度的定量(H)及比率变化曲线(I)。数据为均值±SEM(n=3)。DIC,微分干涉 contrast。(J)人胶质瘤组织切片(患者1)同时用GSHP[绿色(G)/红色(R)通道]和ADRB3抗体(品红色)染色成像。(K)定量分析证实肿瘤区域内各通道强度显著更高。(L)肿瘤区域内荧光比值显著更高。数据为均值±SEM(n=5个视野)。比例尺:10 μm(G)和50 μm(J)。统计显著性采用双尾非配对t检验,P < 0.01;*P < 0.001。
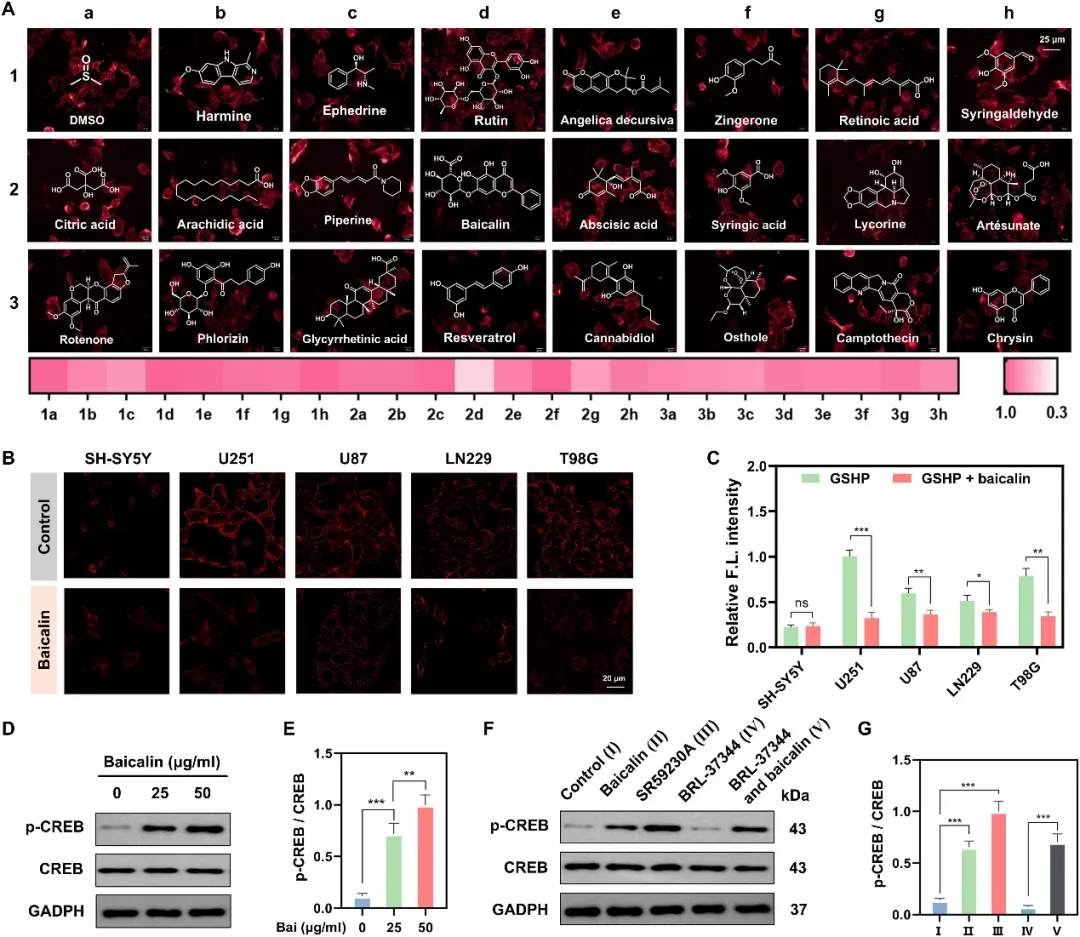
图4. 高通量筛选鉴定黄芩苷为β3-AR抑制剂,可调控多种胶质瘤细胞系中的非经典CREB信号通路。(A)在过表达β3-AR的HEK293T细胞中对天然产物库进行基于荧光的高通量筛选。细胞先用不同化合物(20 μM)预处理,再与GSHP(100 nM)孵育。展示了代表性荧光图像,并叠加了化合物结构。下方的热图总结了相对荧光强度(以DMSO对照组为归一化标准),颜色越浅表示对GSHP信号的抑制作用越强。黄芩苷(位置2d)被鉴定为最强效的抑制剂。比例尺:25 μm。(B)共聚焦图像显示,在多个GBM细胞系(U251、U87、LN229和T98G)中,黄芩苷(25 μg/ml)可竞争性置换GSHP(100 nM)。比例尺:20 μm。(C)对(B)中相对荧光强度的定量分析。U251组(仅GSHP)的信号设为1。(D和E)Western blot(D)及定量分析(E)显示,黄芩苷在U251细胞中以剂量依赖性方式诱导p-CREB。(F和G)Western blot(F)及定量分析(G)证实黄芩苷作为β3-AR抑制剂的作用,其对p-CREB的影响与拮抗剂SR59230A相似,而与激动剂BRL-37344相反。数据以均值±SEM表示(n=3)。统计显著性采用单因素方差分析(ANOVA)进行多重比较,P < 0.05;P < 0.01;P < 0.001。ns,无显著性差异。
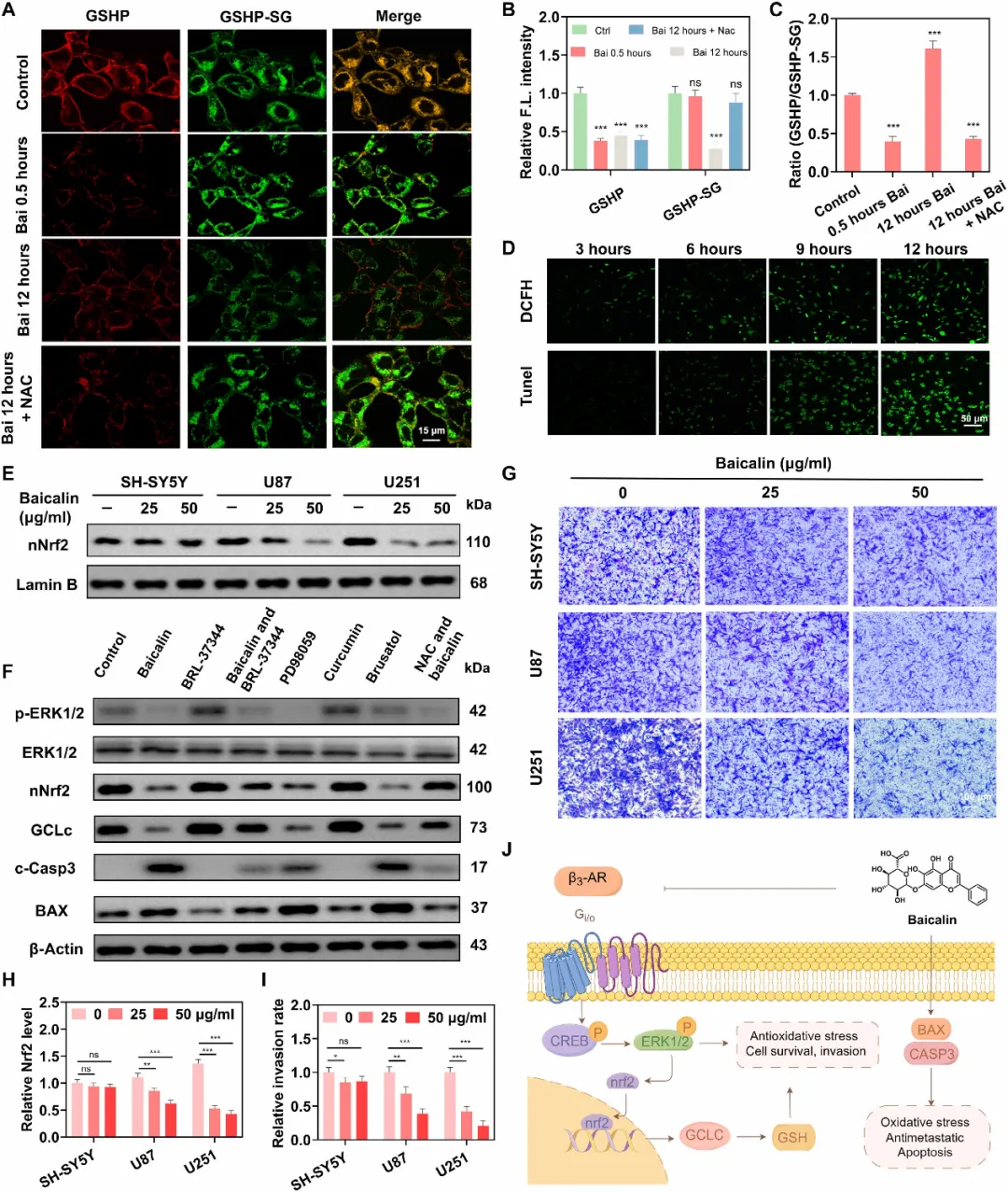
图5. 成像显示黄芩苷通过抑制β3-AR–ERK–Nrf2轴诱导U251 GBM细胞氧化应激并促进细胞凋亡。(A)U251细胞与GSHP(100 nM)孵育后,经黄芩苷(Bai;25 μg/ml)处理不同时间(0.5小时和12小时)或与NAC(5 mM)共处理12小时的共聚焦荧光图像。红色通道(GSHP)显示β3-AR占有率,绿色通道(GSHP-SG)报告局部GSH水平。比例尺:15 μm。(B和C)对(A)中荧光信号的定量分析,显示GSHP和GSHP-SG的相对荧光强度(B)以及GSHP/GSHP-SG比值(C)。(D)经黄芩苷(25 μg/ml)处理不同时间(3、6、9和12小时)后,U251细胞中ROS水平(DCFH-DA染色)和凋亡水平(TUNEL染色)的荧光图像。比例尺:50 μm。(E)Western blot分析显示黄芩苷对不同脑癌细胞系中NRF2水平的影响。Lamin B作为核内参对照。(F)Western blot分析阐明U251细胞经不同药理调节剂处理24小时后的信号通路。处理包括:黄芩苷(25 μg/ml)、β3-AR激动剂BRL-37344(10 μM)、ERK抑制剂PD98059(20 μM)、姜黄素(20 μM)、NRF2抑制剂brusatol(10 μM)以及抗氧化剂NAC(5 mM)。(G)Transwell侵袭实验代表性图像,显示黄芩苷(0、25和50 μg/ml)对不同β3-AR表达水平的细胞系(Sh-SY5Y、U87和U251)的影响。(H)对应(G)的定量分析。(I)(G)中侵袭率的定量分析。(J)使用Figdraw.com制作的黄芩苷作用机制示意图。数据以均值±SEM表示(n=3)。统计显著性采用单因素方差分析(ANOVA)进行多重比较,P < 0.05;P < 0.01;P < 0.001。
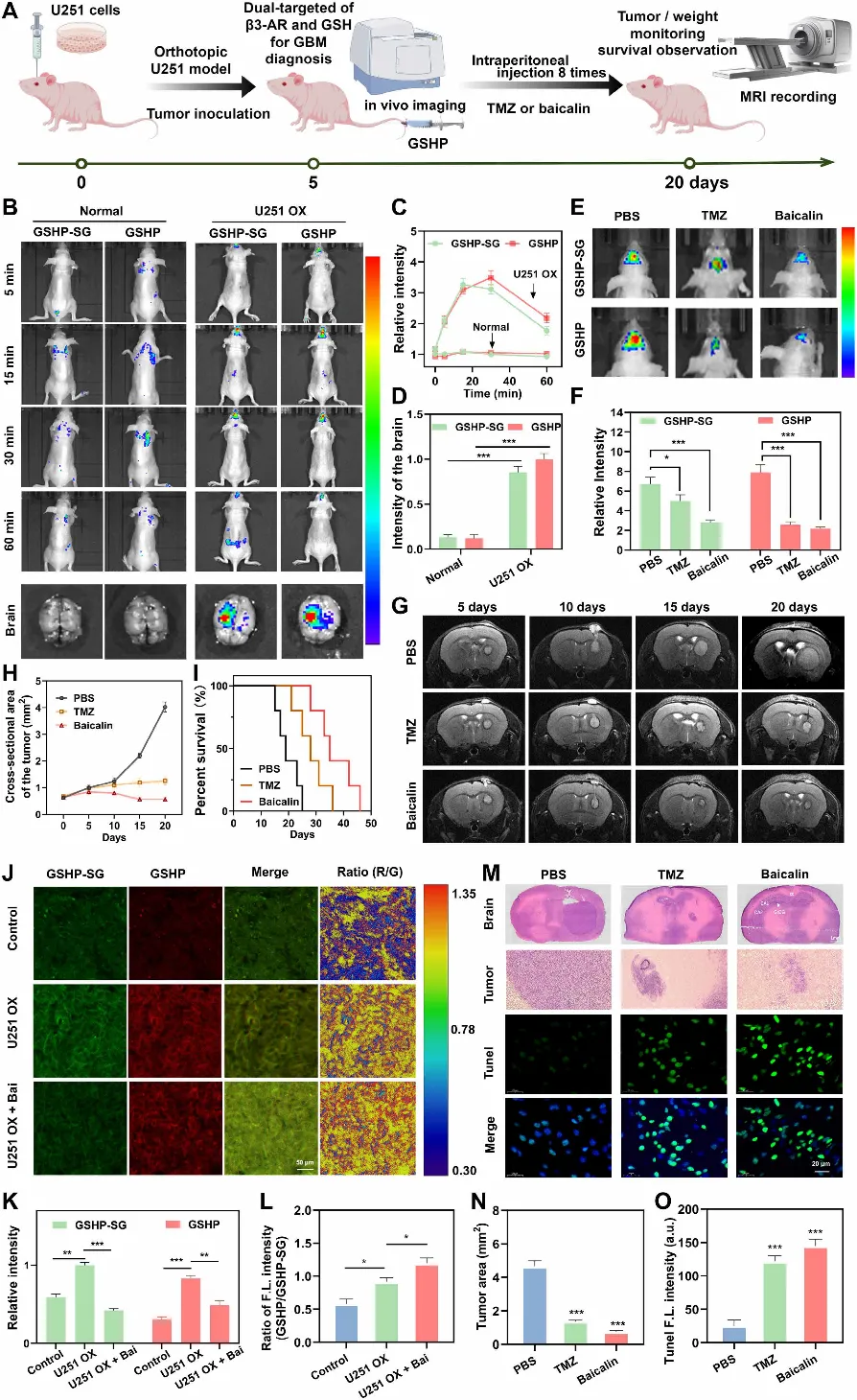
图6. 利用GSHP进行体内双通道成像,用于胶质瘤诊断及黄芩苷疗效的治疗监测。(A)原位U251胶质瘤模型、基于GSHP的双通道成像及治疗监测流程示意图。使用Figdraw.com绘制。(B)正常小鼠与荷原位U251胶质瘤(OX)小鼠体内GSHP-SG和GSHP信号的活体荧光成像。下图:注射后60分钟时的离体脑成像。(C和D)对(B)中GSHP-SG和GSHP信号的定量分析。(E)第20天时不同治疗组小鼠体内GSHP-SG和GSHP信号的活体荧光成像。(F)对(E)中GSHP-SG和GSHP信号的定量分析。(G)代表性MRI图像,显示经PBS、TMZ或黄芩苷治疗的小鼠在20天内肿瘤的进展。(H)基于(G)中MRI图像对肿瘤横截面积的定量分析。(I)不同治疗组U251 OX小鼠的Kaplan-Meier生存曲线。(J)正常小鼠、未治疗的U251 OX小鼠及黄芩苷治疗的U251 OX小鼠脑切片的共聚焦图像。比例尺:50 μm。(K和L)对(J)中GSHP或GSHP-SG信号及其比值的定量分析。(M)各治疗组脑切片的H&E染色(上图)和TUNEL染色(下图)。(N)基于H&E染色切片对肿瘤面积的定量分析。(O)基于TUNEL染色对凋亡率的定量分析。数据以均值±SEM表示(体内实验每组n=5)。
为了促进基于氧化还原的β3-AR生物学研究,并探索靶向β3-AR用于胶质瘤诊断及疗效评估的潜力,本研究报告了一种具有血脑屏障通透性的双功能探针GSHP的创新设计与表征。通过将卡拉洛尔衍生的识别单元与可逆的GSH响应基团相结合,GSHP实现了特异性受体结合和GSH动态的比率检测。这种双功能设计使其能够在从体外系统到体内原位U251 GBM小鼠模型的多种模型中,同时成像β3-AR分布并实时监测氧化还原状态。GSHP不仅能够可视化受体定位,还能捕捉动态的GSH变化,为研究涉及β3-AR的氧化还原生物学提供了强大工具。利用基于GSHP的高通量筛选平台,作者鉴定出黄芩苷是一种竞争性β3-AR配体。通过深入的分子机制研究,作者进一步揭示黄芩苷是β3-AR的天然抑制剂。它破坏β3-AR–ERK–Nrf2信号级联,导致Nrf2的核积聚减少,并下调其下游靶点GCLc。抑制该通路可增强氧化应激,并促进U251胶质瘤细胞凋亡。功能研究证实,黄芩苷显著抑制过表达β3-AR的U251 GBM细胞的侵袭能力,提示调控β3-AR在U251胶质瘤治疗中具有治疗潜力。
除了分子筛选功能外,GSHP在诊断和治疗评估方面也展现出应用前景。对多种脑肿瘤细胞和人胶质瘤组织的成像表明,GSHP有潜力基于β3-AR表达水平和氧化还原状态的差异,区分癌细胞/组织与正常细胞/组织。关键在于,在U251胶质瘤原位小鼠模型中的体内成像证实,GSHP能有效穿过血脑屏障,富集于肿瘤部位,并实现用于肿瘤可视化和治疗反应监测的双通道成像。利用该平台,作者证实经GSHP筛选出的黄芩苷可抑制U251胶质瘤进展并延长生存期,其疗效与GSHP检测到的肿瘤氧化应激增强呈正相关。尽管前景广阔,但本研究存在局限性。大多数发现源自U251细胞系和单一原位小鼠模型,因此需要在多种胶质瘤亚型和临床相关模型中进行未来验证。此外,尽管β3-AR–ERK–Nrf2通路已被确定为本研究的关键调控机制,但仍需进一步研究以探索黄芩苷的其他信号通路或脱靶效应。未来的工作应致力于优化GSHP的药代动力学和稳定性,并将其应用扩展到临床前成像、药物发现和疗效评估中,从而推动黄芩苷或相关β3-AR调节剂进入药物研发管线。总之,GSHP代表了一种连接分子成像、氧化还原生物学和药物发现/疗效评估的强大化学工具,为U251 GBM中靶向β3-AR的诊断和治疗提供了新途径。GSHP成像与靶向β3-AR治疗策略的整合,有望在未来应用于胶质瘤及更广泛的生物医学领域的精准诊断、治疗监测和机制探索中。
文献链接:10.1126/sciadv.aed5337
点击“阅读原文”直达上述文章