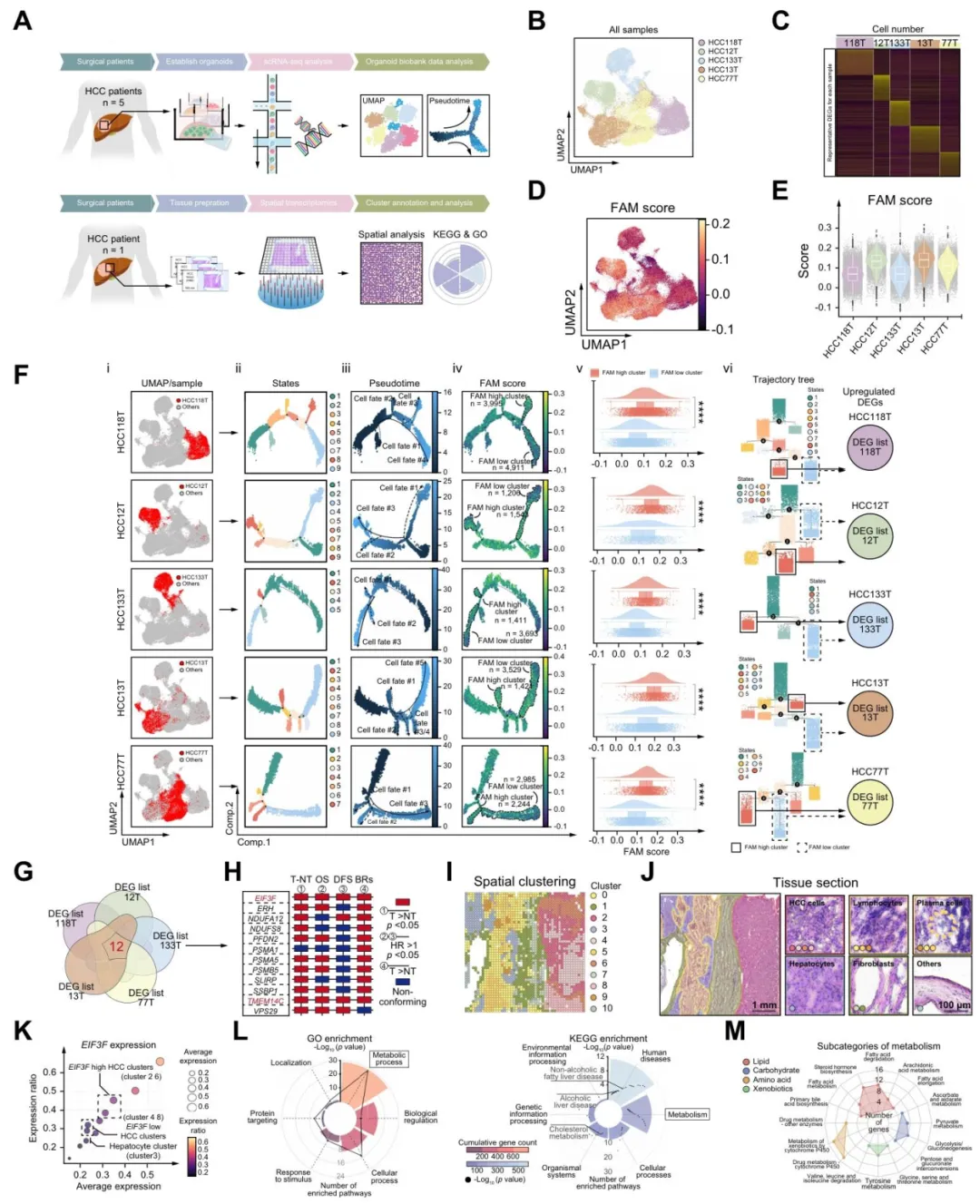
团队建立了5个病人来源的肝癌类器官,做单细胞测序后计算脂肪酸代谢评分。高低评分组做拟时序分析,找差异基因,再结合TCGA生存数据,最后锁定EIF3F和TMEM14C【图1A-G】。
进一步在90对临床样本和空间转录组中验证,eIF3f在肝癌区域明显高表达,并且差异基因富集在脂代谢通路【图1H-M】。
图1:结合类器官单细胞RNA测序与大数据库分析,筛选在肝细胞癌中同时调控脂肪酸代谢(FAM)与肿瘤恶性表型的关键基因
复现思路:类器官数据不好拿,但TCGA+空间转录组+脂肪酸代谢基因集(MsigDB)完全可以在线做,重点是拟时序和生存筛选的逻辑链。
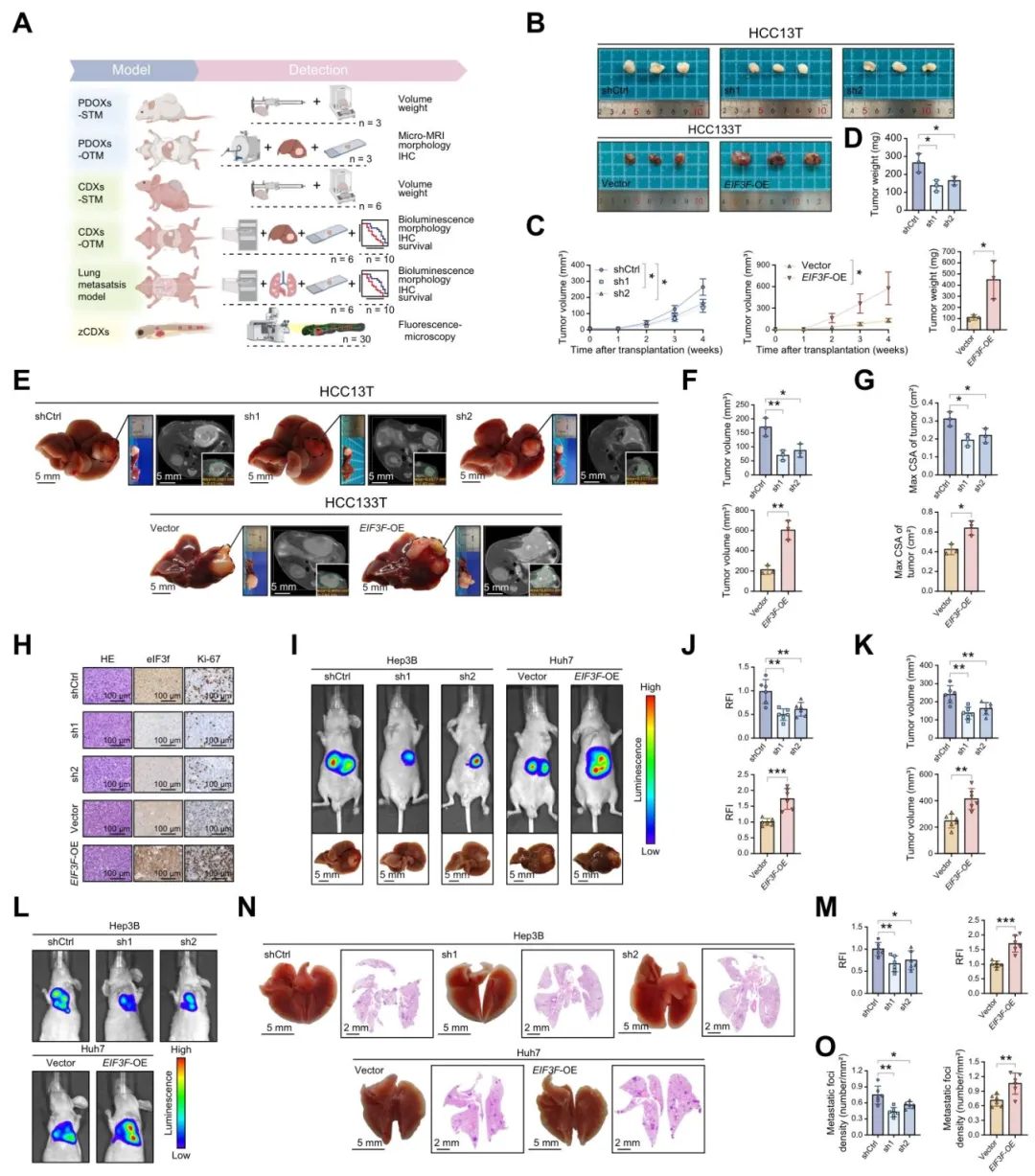
敲低eIF3f后,类器官和小鼠皮下、原位移植瘤体积和重量显著下降,过表达则相反【图2B-G】。Ki67染色也证实增殖能力被明显改变【图2H】。
尾静脉肺转移模型和生存曲线也一致:敲低组转移少、活得更久【图2L-O】。
图2:eIF3f表达上调在体内、体外实验中促进肝细胞癌恶性进展
复现思路:体内实验不好做,但我们可以用公共数据验证eIF3f与增殖、转移相关基因集的相关性,作为旁证。
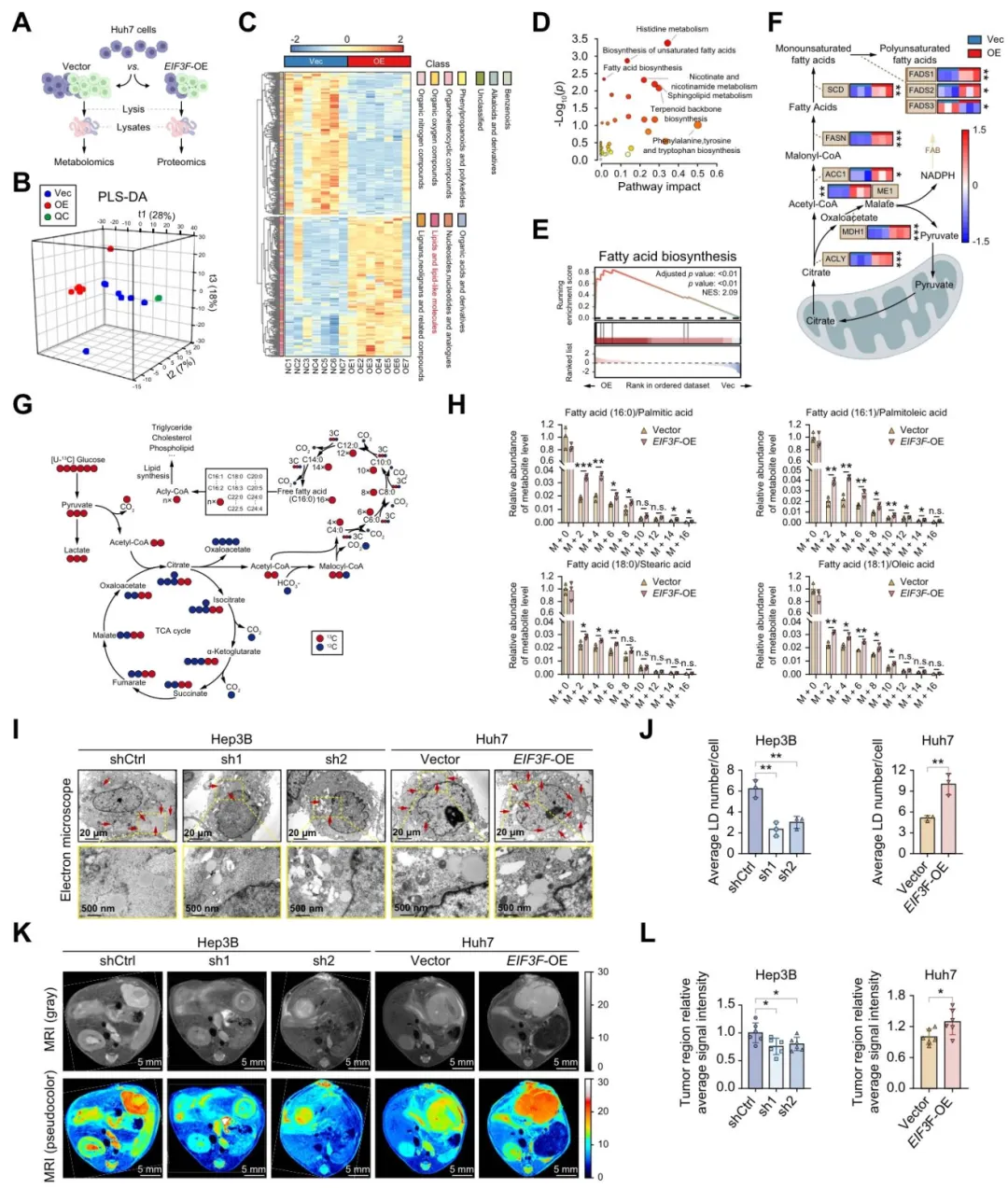
代谢组显示,eIF3f过表达组脂质类代谢物显著上调,尤其是棕榈酰辅酶A、硬脂酸等【图3C-E】。
13C-葡萄糖标记实验证实,eIF3f过表达让葡萄糖更高效地转化为长链脂肪酸【图3G-H】。
电镜和Bodipy染色也看到脂滴数量和脂质含量明显增加【图3I-L】。
图3:eIF3f促进肝细胞癌的脂肪酸生物合成(FAB)与胞内脂质蓄积
复现思路:代谢通量不好做,但我们可以用单细胞数据看eIF3f与脂肪酸合成酶(FASN、ACLY等)的共表达。
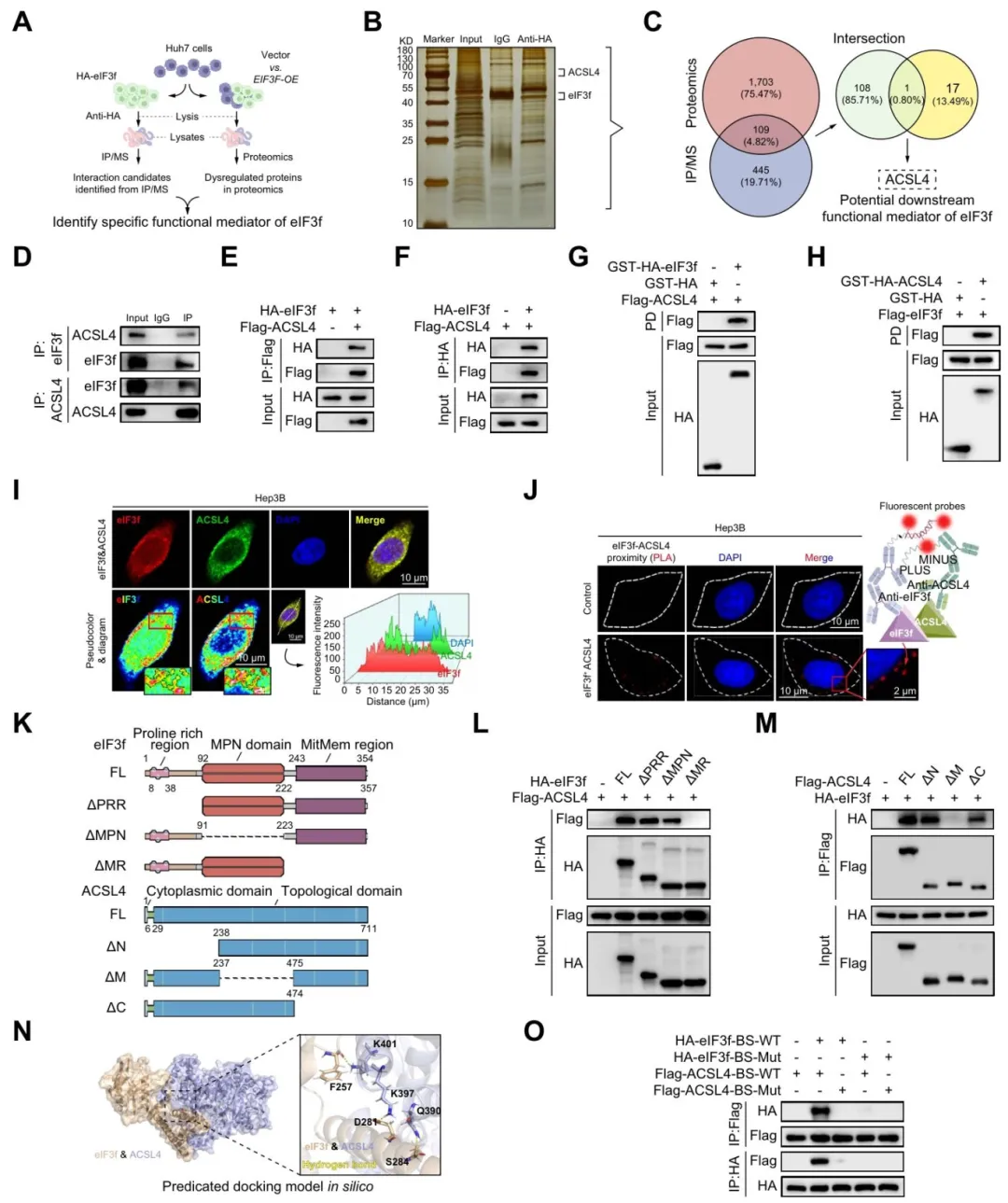
IP-MS+蛋白组+FAB相关基因集取交集,筛出ACSL4作为候选靶蛋白【图4A-C】。Co-IP、GST pull-down和PLA实验都证实两者直接结合【图4D-J】。
截短体实验锁定eIF3f的MIR区(223-357位)和ACSL4的中央区(238-474位)负责结合【图4K-M】。分子对接和点突变验证了F257、S267、K401等关键结合位点【图4N-O】。
图4:eIF3f可特异性结合脂肪酸合成关键分子ACSL4
复现思路:IP-MS数据没有的话,可以用String或BioGRID预测互作,再用TCGA做eIF3f和ACSL4的表达相关性。
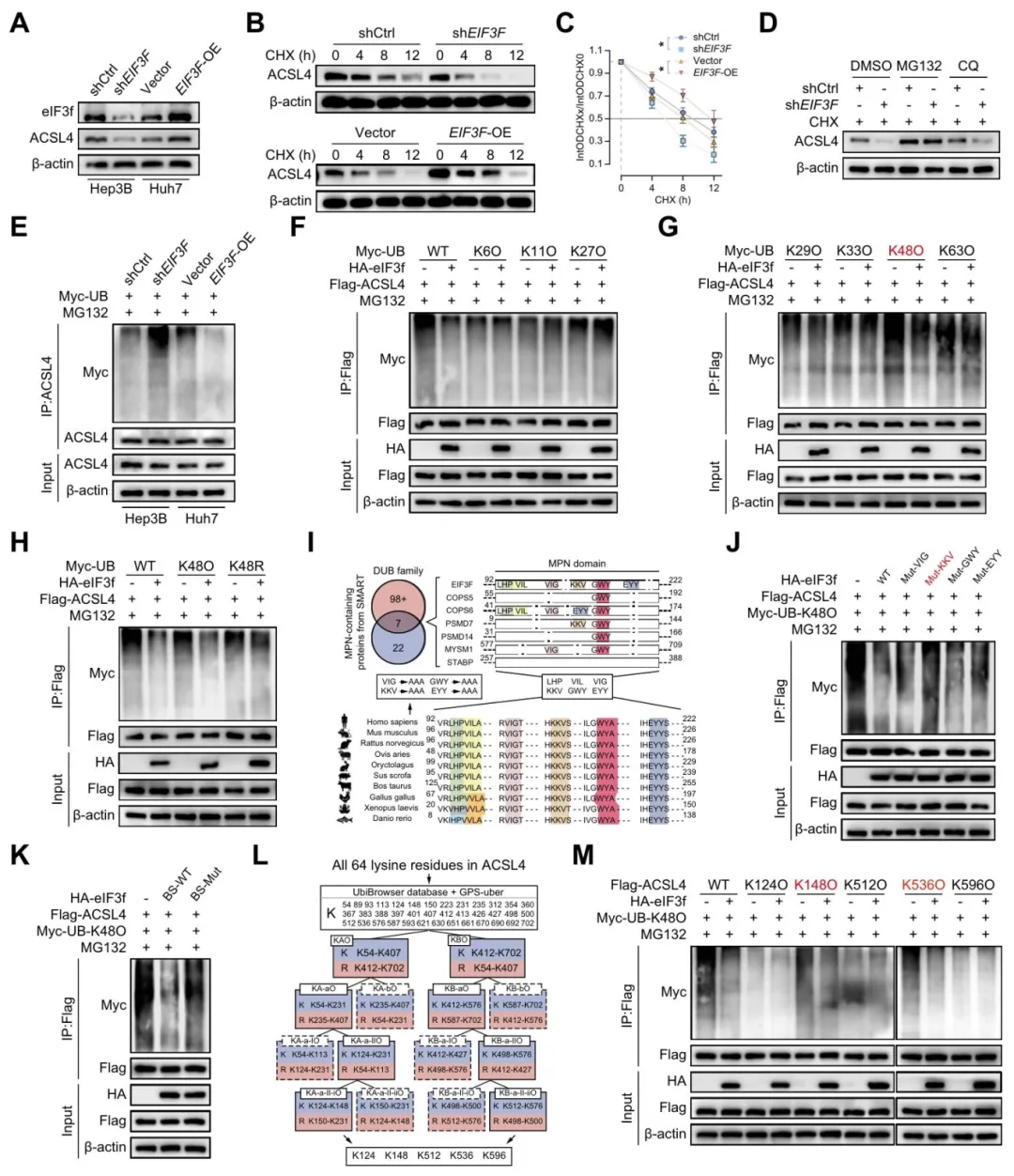
放线菌酮实验显示eIF3f延长ACSL4蛋白半衰期【图5B-C】。MG132可逆转eIF3f敲低导致的ACSL4下降,说明是蛋白酶体途径【图5D】。
eIF3f过表达显著减少ACSL4的K48泛素化,而K48R突变体消除这一效应【图5E-H】。进一步找到eIF3f的MPN结构域中KKV(161-163位)是关键活性位点【图5I-J】。
ACSL4的K148和K536是泛素化的主要修饰位点【图5L-M】。
图5:eIF3f抑制K48位泛素化介导的ACSL4蛋白酶体途径降解
复现思路:泛素化位点可以拿UbiBrowser或GPS-Uber预测,eIF3f的DUB活性可用已知结构域注释推断。
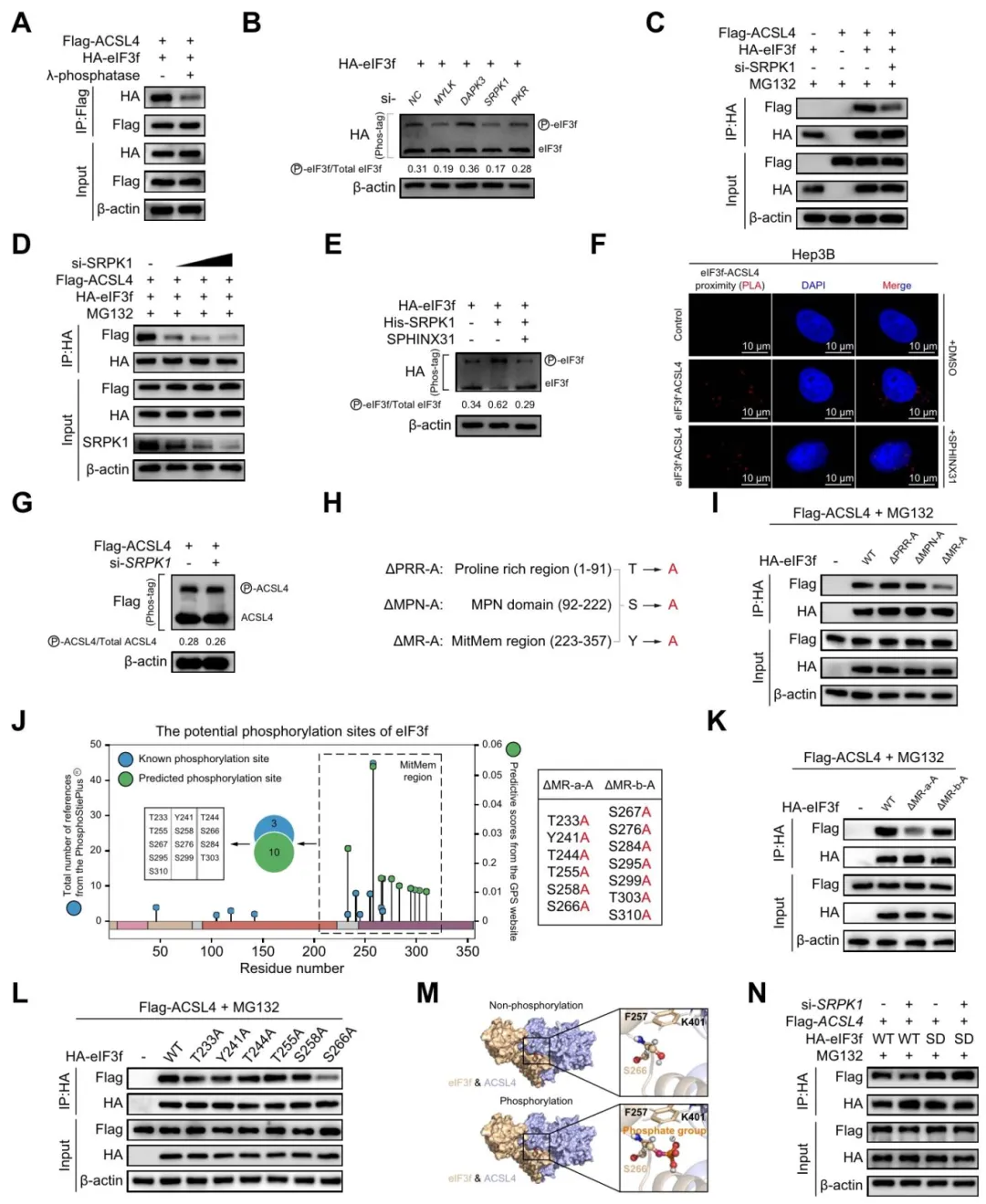
λ-磷酸酶处理后,eIF3f-ACSL4结合显著减弱【图6A】。筛选多个激酶,发现SRPK1敲低降低eIF3f磷酸化水平,并减弱两者结合【图6B-D】。
SPHINX31(SRPK1抑制剂)同样阻断结合【图6E-F】。点突变筛选确认Ser266是eIF3f上受SRPK1磷酸化的关键位点【图6J-N】。
图6:SRPK1介导eIF3f第266位丝氨酸磷酸化,促进eIF3f与ACSL4相互结合
复现思路:磷酸化位点看PhosphoSitePlus,激酶-底物关系可参考Kinase enrichment分析。
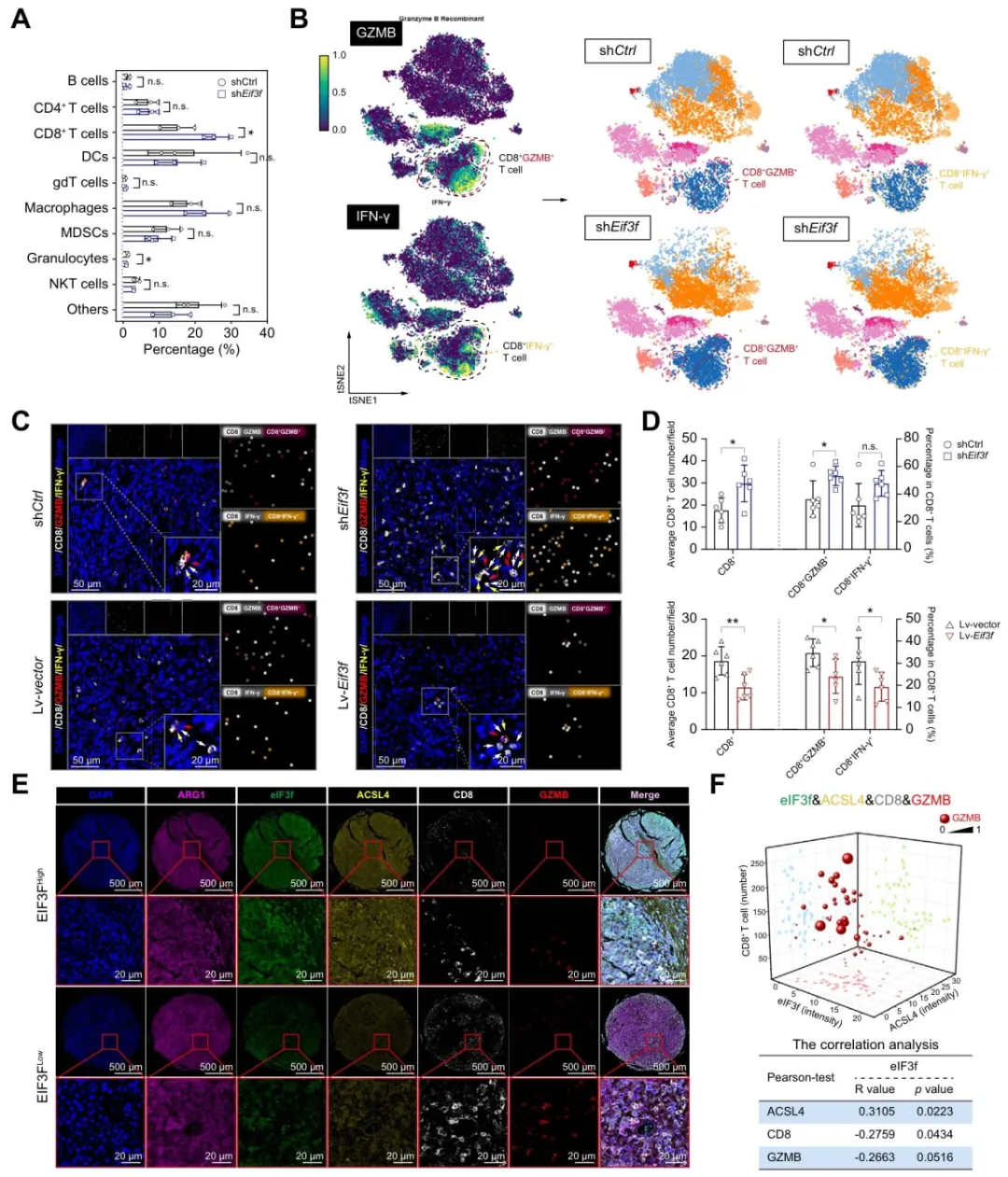
免疫缺陷鼠和免疫正常鼠对比发现,敲低eIF3f在免疫正常鼠中抑瘤效果更明显,提示免疫参与【图S21C】。
质谱流式显示,敲低eIF3f后CD8+ T细胞比例上升,IFN-γ和GZMB也升高【图7A-B】。
CD8抗体清除实验逆转了敲低eIF3f的抑瘤效果【图S23】。临床组织芯片也证实eIF3f高表达与CD8+ T细胞和GZMB水平负相关【图7E-F】。
图7:eIF3f高表达与肝细胞癌组织中CD8⁺T细胞浸润减少相关
复现思路:免疫浸润可以直接用TIMER或xCell基于TCGA做,重点看eIF3f和CD8A、GZMB、IFNG的相关性。
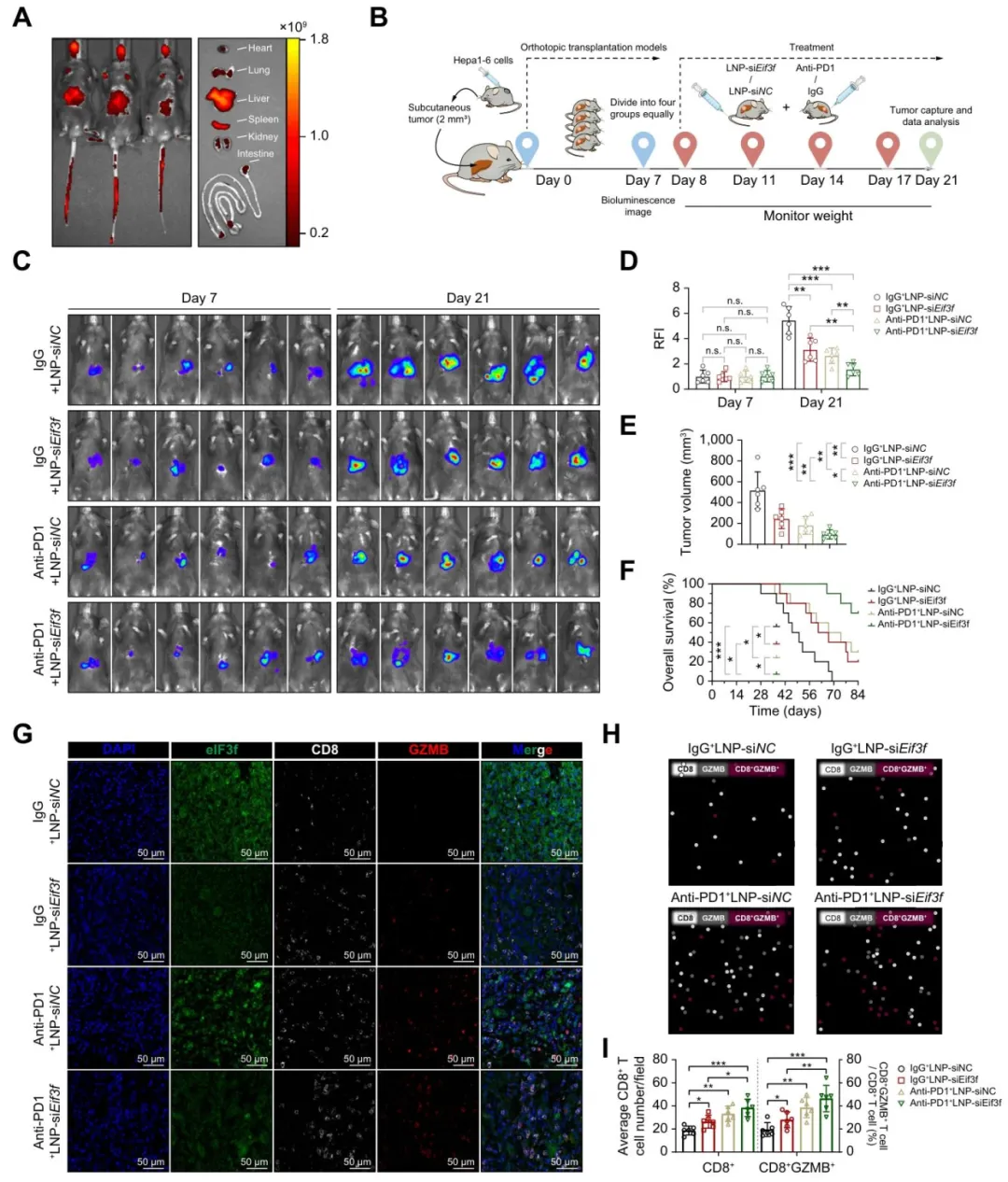
用脂质纳米颗粒包裹siEIF3F,能有效递送到肝脏【图8A-B】。单用LNP-siEIF3F或抗PD-1都能抑瘤,但两者联用效果最好,肿瘤体积最小、生存期最长【图8C-F】。
多色免疫组化也证实联合治疗组CD8+ T细胞和GZMB+ CD8+ T细胞比例最高【图8G-I】。
图8:靶向eIF3f可减轻小鼠肿瘤负荷并提升抗PD‑1疗法的抗肿瘤效果
复现思路:这里主要是临床前实验,生信层面可做eIF3f与免疫检查点(PD-1、PD-L1、CTLA4)共表达及预后分层分析。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
