mBio|南京农业大学动物医学院最新研究:猪流行性腹泻病毒变异株免疫逃逸新机制-S2亚基894–993氨基酸区成关键调控区域
猪场里仔猪突然出现剧烈水样腹泻、迅速脱水死亡的场景,近年来已成为许多养殖户最头疼的问题。自2010年以来,猪流行性腹泻病毒(PEDV)变异株在全球范围内广泛流行,导致数百万仔猪死亡,给养猪业造成巨大经济损失。传统基于经典株(GI型)研制的疫苗对这些变异株(GII型)保护效果显著下降,其背后的分子机制长期不明。2026年3月23日,国际期刊mBio在线发表了一篇题为《Evasion of humoral immune responses by a key mutational region of the S2 subunit in PEDV variants》的研究论文,南京农业大学动物医学院牵头,联合江苏农业科学院等单位的研究团队,通过交叉中和试验、重组病毒构建、仔猪攻毒保护试验以及结构生物学分析,明确S2亚基894–993氨基酸区是PEDV变异株逃避体液免疫、增强毒力和调控膜融合的关键区域。这一发现为理解变异株的致病机制提供了新证据,也为研发更有效的防控策略奠定了基础。
变异株与经典株的免疫差异:交叉中和试验揭示“逃逸”现象
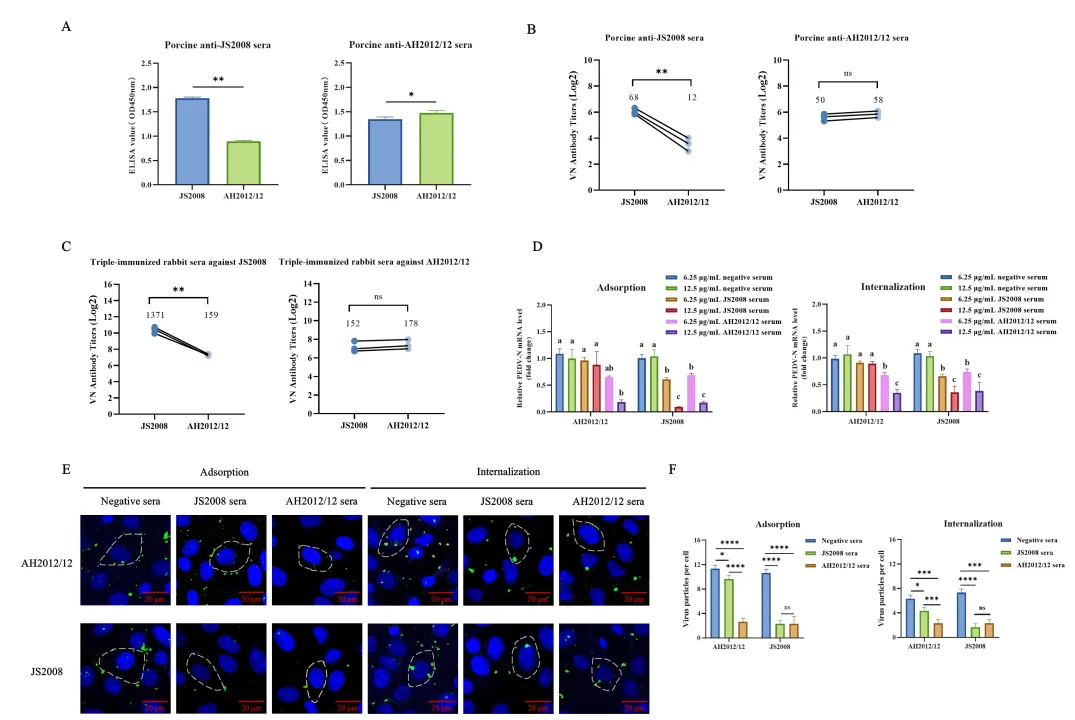
PEDV是一种包膜的单股正链RNA病毒,其棘突蛋白(S蛋白)是病毒附着、进入宿主细胞并介导膜融合的关键结构。S蛋白由S1亚基(负责受体识别)和S2亚基(负责膜融合)组成。研究团队首先制备了针对经典GI株JS2008和变异GII株AH2012/12的猪源和兔源高免血清,并进行了系统的交叉中和试验。
结果显示,抗JS2008血清对同源JS2008病毒的中和效价约为1:68,但对异源AH2012/12病毒的中和效价显著降低至1:12左右;而抗AH2012/12血清对两种病毒均表现出较强的中和活性。此外,抗JS2008血清仅能有效抑制同源病毒的吸附和内化阶段,对变异株则几乎无抑制作用;抗AH2012/12血清则在两个阶段均能有效抑制两种病毒。这些数据表明,PEDV变异株在体液免疫层面存在明显的逃逸能力,经典株诱导的抗体难以有效中和变异株。
这一现象与临床观察高度一致:免疫过经典株疫苗的猪群对变异株仍高度易感,提示S蛋白上的特定突变可能是导致免疫逃逸的主要原因。
重组病毒筛选:锁定S2亚基894–993氨基酸区为免疫逃逸核心
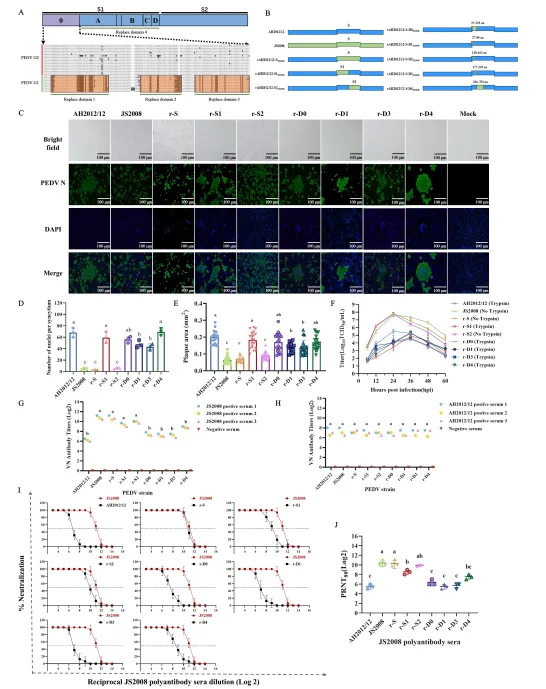
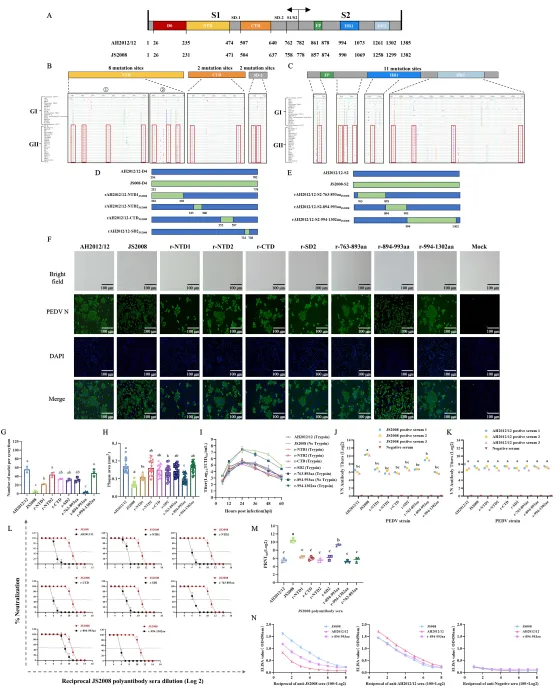
为精准定位关键突变区域,研究团队以AH2012/12为骨架,利用CRISPR/Cas9基因编辑技术,系统构建了一系列S基因替换的重组病毒,包括全S、S1、S2、D0区以及D0区进一步细分的片段。随后又针对D4区和S2亚基进行小片段替换,共拯救出多个稳定复制的重组病毒。
交叉中和试验结果显示,只有将S2亚基或894–993氨基酸区替换为JS2008序列的重组病毒(如r-S2和r-894-993aa),才能被抗JS2008血清高效中和;而其他重组病毒仍保持对经典株血清的强抵抗力。空斑减少中和试验(PRNT)和ELISA结合试验进一步证实,r-894-993aa与抗JS2008血清的结合活性显著高于亲本AH2012/12。这表明S2亚基894–993氨基酸区是变异株逃避经典株诱导中和抗体的关键决定因素。
值得注意的是,此前研究多关注S1亚基的N端区域(如D0区),认为其是主要抗原表位。而该研究首次证明S2亚基内部的这一特定区域在免疫逃逸中发挥核心作用,且该区域的氨基酸突变在GII亚型株中高度保守,提示其可能是病毒在疫苗选择压力下适应性进化的结果。
仔猪体内试验:894–993区直接影响病毒毒力
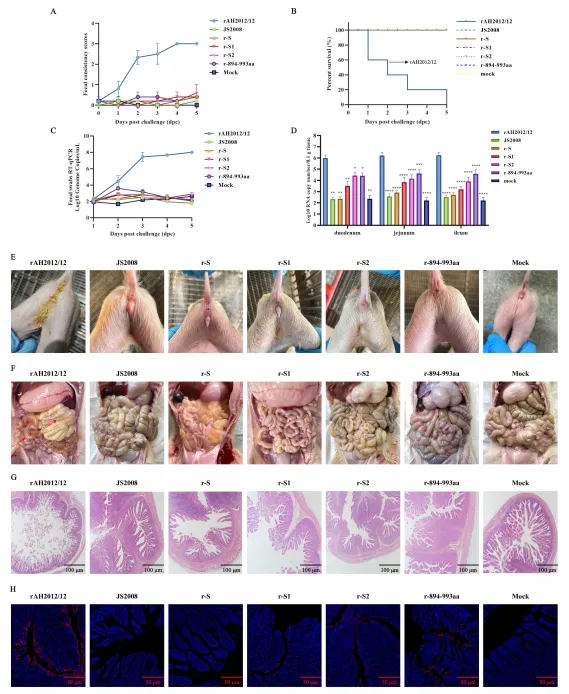
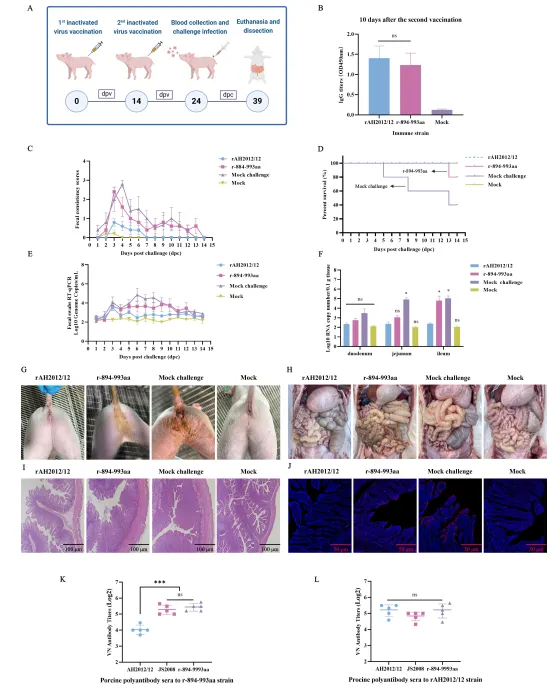
体外结果需经体内验证。研究团队选取3日龄PEDV阴性仔猪,分别口服攻毒rAH2012/12、JS2008、r-S、r-S1、r-S2以及r-894-993aa等毒株,并设置空白对照组。
攻毒后观察显示,rAH2012/12组仔猪1天后即出现水样腹泻,4–5天症状加重,病毒排出量在2–5天逐步升高至10^8拷贝/mL,死亡率较高;肠道剖检可见小肠膨胀、变薄,组织病理显示肠绒毛严重萎缩、坏死。而r-894-993aa组仅出现轻度腹泻,病毒排出量显著降低,其他重组病毒组临床症状不明显。病毒载量检测和免疫荧光结果也证实,r-894-993aa感染组肠道病毒复制水平远低于rAH2012/12组。
这些结果表明,894–993氨基酸区不仅是免疫逃逸的关键区域,同时也是决定变异株高毒力的重要结构域。S1和S2亚基均参与毒力调控,但S2亚基内的这一小片段作用尤为突出。
免疫保护试验:进一步验证该区在体内免疫逃逸中的作用
为评估该区域在实际免疫场景中的意义,研究团队制备了rAH2012/12和r-894-993aa灭活疫苗,免疫7日龄仔猪,间隔14天加强免疫,随后用rAH2012/12强毒攻毒。
结果显示,rAH2012/12疫苗组仔猪攻毒后仅出现轻度腹泻,病毒排出量低,肠道组织基本正常;r-894-993aa疫苗组则与未免疫攻毒组相似,出现明显腹泻和高病毒载量,仅存活率略有提升。血清学检测进一步证实,抗r-894-993aa血清对自身和JS2008中和效果良好,但对AH2012/12中和能力较弱;而抗rAH2012/12血清对多种毒株均有较好中和活性。
这一体内试验直接证明,894–993氨基酸区的突变使变异株能够有效逃避经典株疫苗诱导的保护性免疫。
传播模式与结构机制:细胞间传播增强与S蛋白构象变化
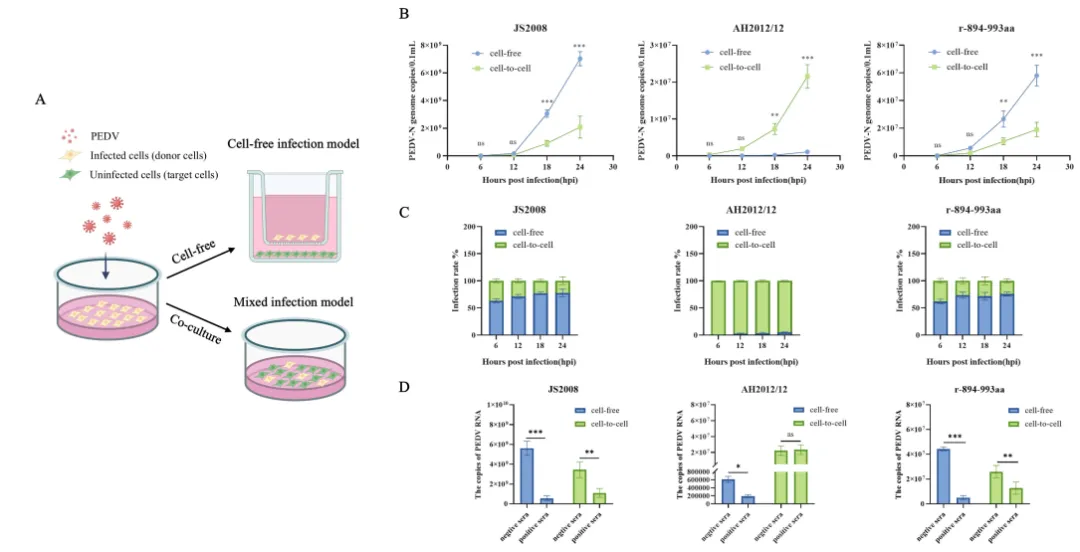
研究团队进一步探究了该区域的分子机制。首先构建了细胞游离传播和细胞-细胞接触传播模型。结果显示,AH2012/12主要依赖细胞-细胞接触方式传播,效率高达95%左右,可有效躲避免疫血清;JS2008和r-894-993aa则以细胞游离传播为主,易被血清抑制。将894–993区替换为经典序列后,重组病毒失去细胞融合能力,传播模式转向细胞游离,抗体抑制效果显著增强。
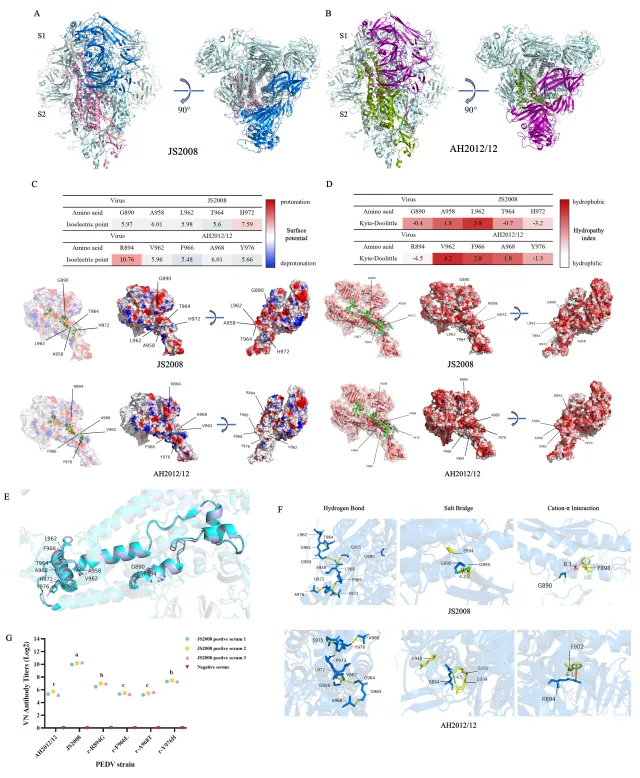
结构生物学分析采用SWISS-MODEL对JS2008和AH2012/12的S蛋白进行同源建模。比较894–993区(对应连接区CR)发现,变异株突变改变了表面静电势(例如R894引入正电荷)、增加了疏水性,并形成新的盐桥和阳离子-π相互作用,使S蛋白构象更刚性、更易向内折叠,从而掩盖潜在中和表位。单氨基酸突变病毒试验显示,894和976位点的突变对免疫逃逸贡献较为突出。
这些机制解释了变异株如何在早期入侵阶段抵抗抗体,同时通过增强细胞间传播进一步实现免疫逃逸。
研究意义与展望:为PEDV防控提供理论支撑
该研究首次系统阐明了S2亚基894–993氨基酸区在PEDV变异株生物学特性中的核心作用,填补了S2亚基功能研究的空白。此前虽然已知S2亚基参与膜融合和部分中和表位,但该特定区域的调控作用尚未被报道。
这一发现不仅揭示了经典疫苗保护失效的分子基础,还为新型疫苗设计提供了潜在靶点。例如,针对该区域开发亚单位疫苗、广谱中和单克隆抗体,或设计小分子抑制剂,均有可能提升对GII变异株的保护效力。同时,研究也提示,在疫苗研发中需兼顾S1和S2亚基的协同作用,以实现更广谱的免疫保护。
PEDV防控仍面临诸多挑战,但随着对病毒关键功能域的深入解析,未来有望开发出更安全高效的疫苗和抗病毒药物,降低养猪业损失。该研究也为冠状病毒家族的跨种传播预警和通用疫苗研发提供了重要参考
原文链接:https://doi.org/10.1128/mbio.00026-26
注:文中插图源于mBio,欢迎关注交流。
本文为原创编译,仅代表对原文内容的整理与解读,原文版权归原作者及出版方所有。本文仅用于学术交流与分享,旨在促进研究与学习。如内容存在错误或涉及侵权,请通过后台私信联系我们进行更正或删除。(当前本公众号的原创文章均为对原文的整理与解读,原文版权归原作者及出版方所有)