南京农大张正光教授团队发表Nature Plants揭示稻瘟病治病的叶绿体蛋白降解机制并利用水稻自身叶绿体Clp的防御机制提升抗性
Nature Plants | 南京农业大学张正光团队2026年4月13日在国际植物生物学杂志Nature plants 发表题为:A fungal nuclease effector subverts the chloroplast genome and triggers cell death to promote infection的研究论文(https://doi.org/10.1038/s41477-026-02276-x)
研究以稻瘟病菌(Magnaporthe oryzae)为对象,从侵染中后期上调的 298 个效应蛋白中筛选出6 个坏死营养型效应子(MoNee1–MoNee6),其中MoNee6作为核酸酶靶向水稻叶绿体并降解叶绿体 DNA、诱导宿主细胞死亡,驱动病菌从活体营养向坏死营养转型;MoNee6 会被水稻叶绿体Clp 蛋白酶降解,但病菌会抑制叶绿体编码的OsClpP1表达削弱该防御;研究通过将OsClpP1 融合叶绿体转运肽实现核编码表达,显著增强水稻对稻瘟病的抗性,且不影响农艺性状,揭示了真菌效应子劫持叶绿体基因组的新致病机制与抗病改造新策略。该研究关注目标蛋白叶绿体定位对其功能的重要性,对叶绿体定位的蛋白的研究具有非常重大的指导意义,也对稻瘟病的防治提供解题思路。
一、研究背景与科学问题
稻瘟病菌是典型半活体营养真菌,需完成活体营养→坏死营养转型才能获取营养、完成致病与产孢,该过程是致病关键,但调控分子机制尚不明确。
科学问题:
稻瘟病菌是否分泌坏死营养型效应子(NEEs)驱动营养型转换?该研究的思维导图:
三、详细总结
1. 研究背景与目的
稻瘟病菌是半活体营养真菌,需从活体营养切换至死体营养获取营养、引发病害,但其营养转换的分子机制尚不明确;本研究旨在筛选病菌侵染后期上调的坏死性效应子,解析其作用机制,并开发水稻抗病改造策略。
2. 效应子筛选与鉴定
- 筛选范围:298 个侵染中后期上调的候选效应蛋白
- 鉴定结果:获得6 个坏死性效应子(MoNee1–MoNee6)
- 功能验证:
- 敲除 MoNee1–6 均降低病菌毒力,ΔMoNee6毒力缺陷最显著
- MoNee6Δsp 在水稻原生质体中诱导最高水平细胞死亡
- 该类效应子在半活体 / 死体营养真菌中保守,活体营养真菌中缺失
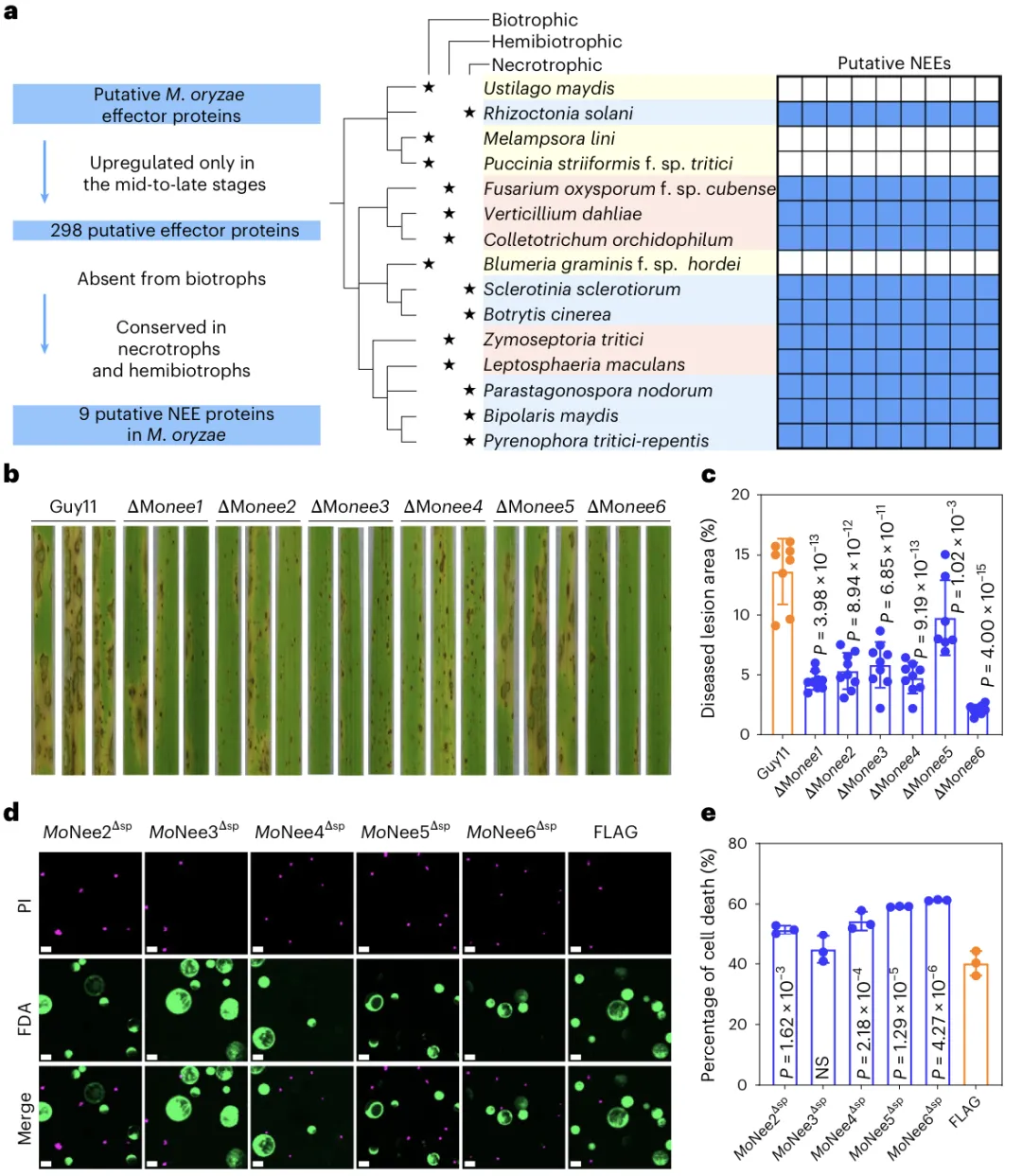
a.系统发育树展示298 个在稻瘟病菌侵染中后期上调的候选效应蛋白,在活体营养型、半活体营养型、死体营养型真菌中的分布情况;重点突出:只在半活体 + 死体营养真菌中保守在活体营养真菌中缺失的坏死性效应子b. 水稻叶片发病表型分别用野生型菌株Guy11与MoNee1–MoNee6 敲除突变体(ΔMonee1–ΔMonee6)侵染水稻叶片,观察并对比病斑症状。c. 病斑面积统计d. 水稻原生质体共聚焦显微成像,对野生型、瞬时表达MoNee2Δsp–MoNee6Δsp及 FLAG 蛋白的水稻原生质体进行FDA/PI 双染色:e. 原生质体细胞死亡率统计
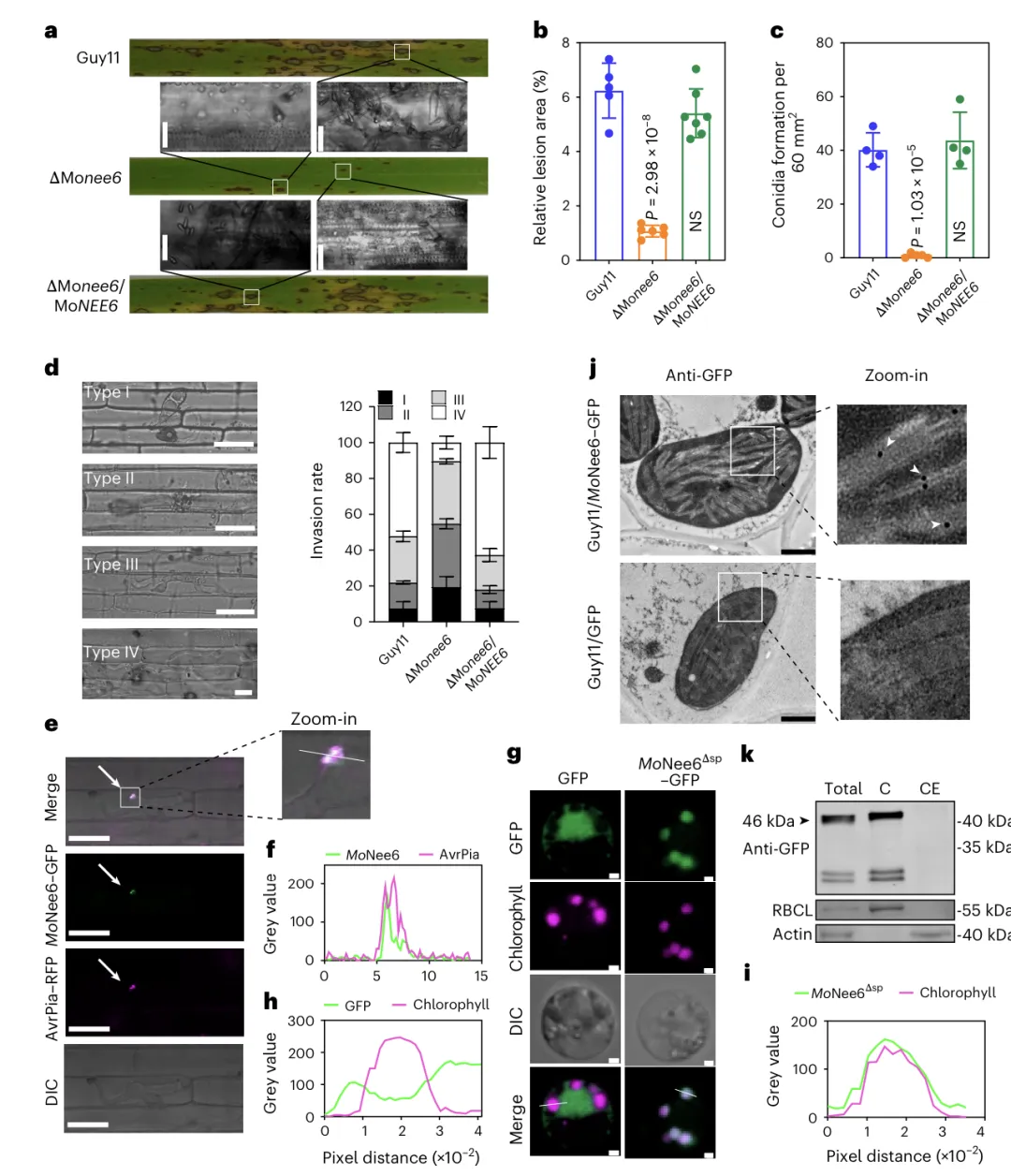
3.1 亚细胞定位,侵染时定位于活体营养界面复合体(BIC),分泌后靶向水稻叶绿体,免疫电镜与蛋白组分检测均验证•定位于叶绿体的 MoNee6 对稻瘟病菌的侵染至关重要 a 野生型、敲除株与回补株侵染水稻后,病斑大小和产孢量均证明 MoNee6 对毒力必需。b–c 敲除 MoNee6 使病斑面积与产孢量显著下降,回补后可恢复。d MoNee6 是病菌在侵染后期向周围细胞扩展的关键因子。e–f MoNee6 在侵染时定位于活体营养界面复合体(BIC)。g–i MoNee6 进入水稻细胞后特异性靶向叶绿体。j 免疫电镜直接证实 MoNee6 存在于叶绿体内部。k 蛋白组分检测进一步验证 MoNee6 富集在叶绿体中。
实验结果证明,叶绿体定位的 MoNee6 是稻瘟病菌关键毒力因子MoNee6 在侵染时定位于 BIC,进入宿主细胞后靶向叶绿体;敲除该基因会显著降低病菌毒力与侵染扩展能力。
3.2 分子功能
结构:含DUF1524 核酸酶结构域,HxxP 基序为活性位点功能:作为非特异性核酸酶,优先降解叶绿体 DNA(cpDNA)下游效应:抑制叶绿体基因转录、激活叶绿体逆行信号、直接诱导宿主细胞死亡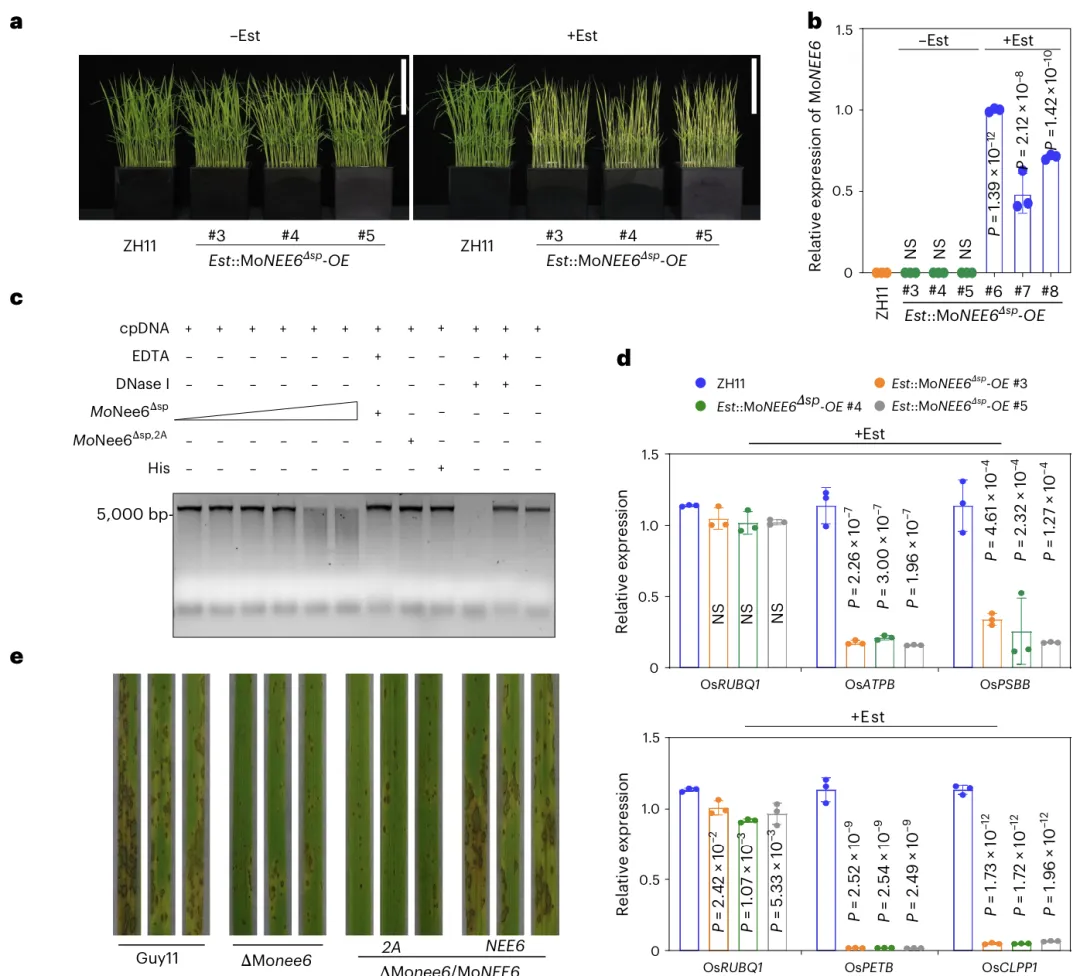
MoNee6 在水稻细胞内降解叶绿体 DNA;a 诱导表达 MoNee6Δsp 使水稻植株黄化、生长受抑。b qRT-PCR 证实 Est 可成功诱导 MoNEE6 高表达。c 体外实验证明 MoNee6Δsp 可降解叶绿体 DNA,失活突变体 MoNee6Δsp,2A 无此活性。d 诱导表达 MoNee6 显著抑制叶绿体基因表达,对核基因影响很小。e 野生型、回补株可引发典型病斑,敲除株与点突变回补株致病力显著下降。f 流式细胞术显示,MoNee6 在侵染过程中直接导致叶绿体 DNA 完整性下降。
3.3 作用保守性
MoNee6 同源蛋白在16 属 180 株病原真菌中广泛存在禾谷镰孢、灰葡萄孢等同源物均具核酸酶活性并诱导细胞死亡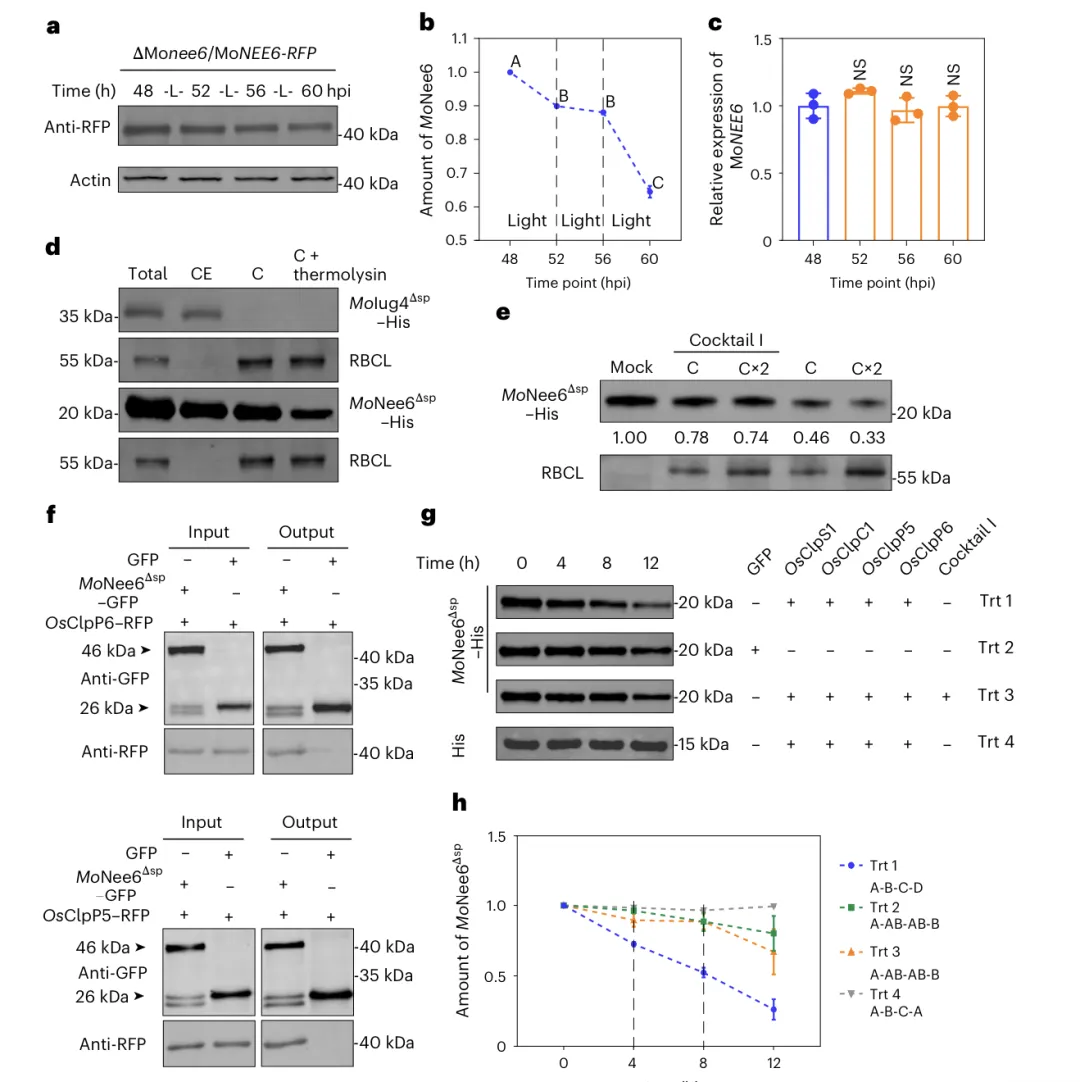
Clp 蛋白酶介导 MoNee6 在水稻叶绿体中的降解; a 光照下,侵染后期 MoNee6–RFP 蛋白水平随时间显著降低。b 灰度定量显示 MoNee6 蛋白在光照下持续降解。c MoNEE6 转录水平无明显变化,降解发生在蛋白水平。d 体外实验证实 MoNee6Δsp 可进入叶绿体内部。e 叶绿体可降解 MoNee6Δsp,蛋白酶抑制剂能减缓该过程。f Co-IP 证明 MoNee6 与 OsClpP5、OsClpP6 直接互作。g–h 完整 Clp 蛋白酶复合体可高效降解 MoNee6。
4. 宿主防御机制:Clp 蛋白酶
4.1 作用方式
MoNee6 在叶绿体中被Clp 蛋白酶复合体降解,且降解具有光依赖性,需完整组装的 Clp 复合体才有降解活性,单一亚基过表达无效4.2 关键亚基 OsClpP1
编码位置:叶绿体基因组编码;病菌干扰:稻瘟病菌通过 MoNee6抑制 OsClpP1 转录,降低 Clp 活性;互作验证:MoNee6 与OsClpP5/6直接互作,OsClpP1 为复合体核心催化亚基
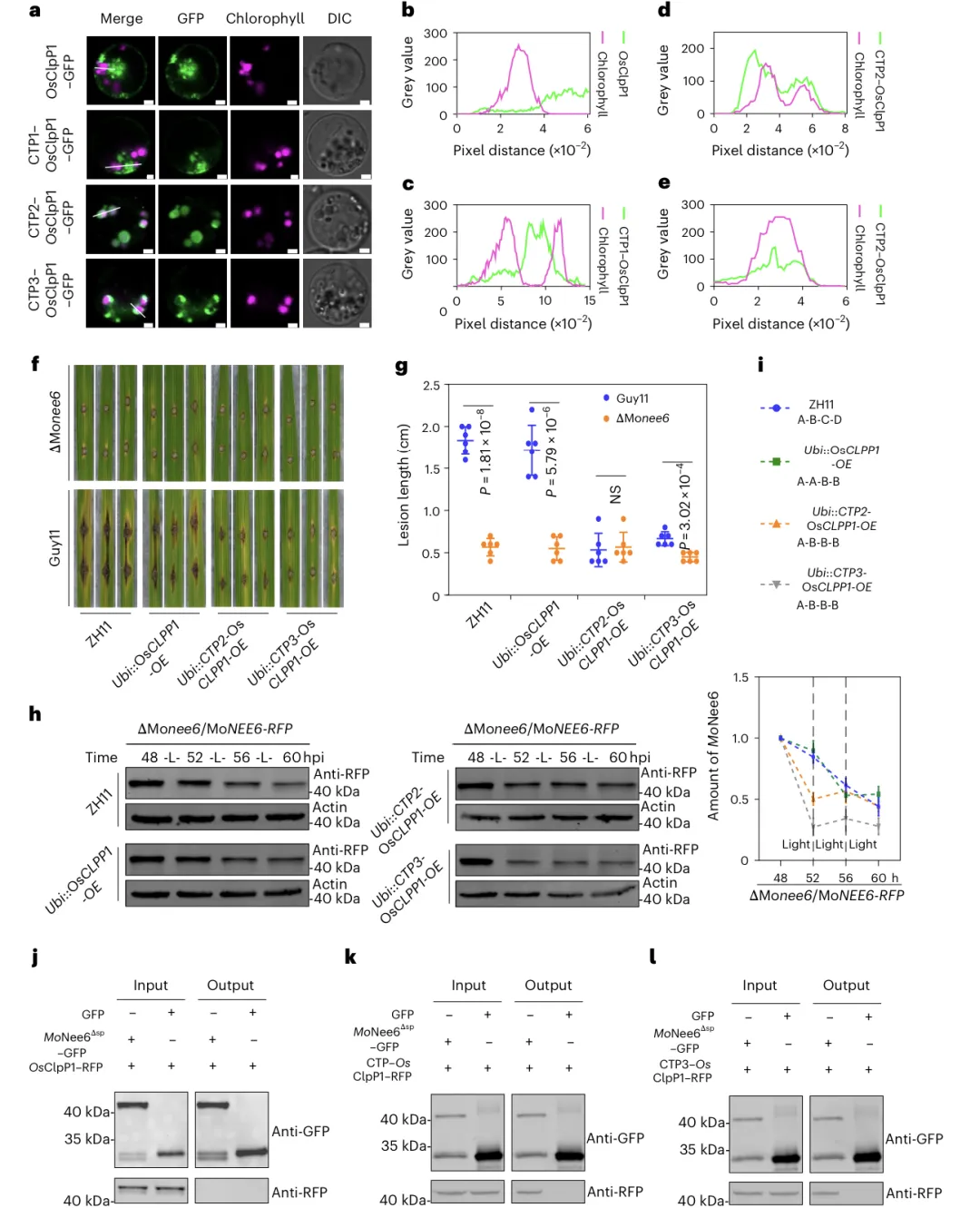
核表达 OsClpP1 增强水稻抗稻瘟病能力; a–e 融合叶绿体转运肽(CTP2/3)可将核编码的 OsClpP1 成功定位到叶绿体。f–g 转 CTP-OsCLPP1 水稻病斑显著减小,抗性明显提升。h–i 改造株中 MoNee6 蛋白被快速降解,含量远低于野生型。j–l 改造后的 CTP-OsClpP1 可与 MoNee6 直接互作,原生 OsClpP1 则不能。
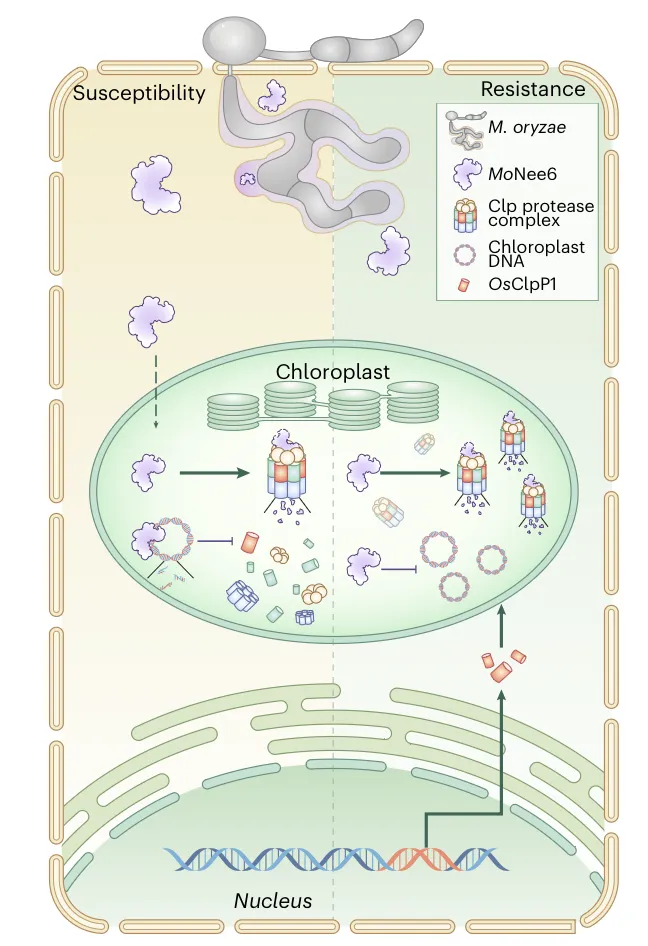
MoNee6 与 Clp 在稻瘟病菌 — 水稻互作中的作用总结:死体营养阶段,MoNee6 进入水稻叶绿体,降解叶绿体 DNA、抑制叶绿体基因转录并引发细胞死亡。部分 MoNee6 会被 Clp 蛋白酶降解,但 MoNee6 造成的叶绿体 DNA 损伤会抑制叶绿体编码亚基OsClpP1的转录,削弱 Clp 活性,使其发挥毒力。工程水稻通过表达核编码的 CTP-OsClpP1 融合蛋白,绕过转录抑制、恢复 Clp 活性、加速 MoNee6 降解,在不影响农艺性状的前提下显著提高抗稻瘟病能力
5. 水稻抗病改造与效果
5.1 改造策略
融合叶绿体转运肽 CTP2/3,实现蛋白精准回运至叶绿体6. 研究结论
首次揭示MoNee6 通过降解叶绿体 DNA 驱动稻瘟病菌营养转换的新机制明确叶绿体 Clp 蛋白酶是植物抵御效应子攻击的重要免疫屏障建立核编码 - 叶绿体靶向改造 OsClpP1的广谱抗病新策略四、关键问题与答案
问题 1:MoNee6 为何是稻瘟病菌致病的关键效应子?
答案:①MoNee6 是6 个坏死性效应子中毒力贡献最强的蛋白,敲除后病菌病斑面积、产孢量、侵染扩散能力均大幅下降;②它是特异性靶向叶绿体的核酸酶,直接降解叶绿体 DNA、抑制叶绿体功能并诱导宿主细胞死亡,是病菌从活体营养转向死体营养的核心驱动因子;③其同源蛋白在多种病原真菌中保守,是病菌通用的致病工具。
问题 2:为何单独过表达 OsClpP5/6 无法提升水稻抗病性?
答案:①Clp 蛋白酶需完整组装的复合体才具备降解 MoNee6 的活性,单一亚基过表达无法组装功能复合体;②OsClpP1 是叶绿体编码的核心催化亚基,稻瘟病菌会特异性抑制其转录,成为 Clp 活性的限速因子;③OsClpP5/6 过表达无法弥补 OsClpP1 的表达抑制,因此无法增强对 MoNee6 的降解效率。
问题 3:核编码 - 叶绿体靶向改造 OsClpP1 的抗病原理是什么?
答案:①原生 OsClpP1 由叶绿体基因组编码,易被 MoNee6 抑制转录;改造后转为核编码,摆脱病菌的转录抑制;②融合CTP2/3 叶绿体转运肽,使核表达的 OsClpP1 精准进入叶绿体,恢复 Clp 蛋白酶复合体的完整活性;③活性提升的 Clp 复合体能快速降解入侵的 MoNee6,阻止叶绿体 DNA 降解与细胞死亡,最终增强水稻对稻瘟病的抗性,且不影响农艺性状。

 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
