南京医科大学 & 徐州医科大学团队2026年发表42.5分新研究:肿瘤氨代谢重编程 Treg,破解免疫治疗耐药新机制
- 2026-05-09 15:02:15

医研新境
课题、实验、培训 全流程设计
欢迎扫码咨询
微信号丨ztsw0925
抖音丨92749311371
导 读

调节性 T 细胞(Tregs)在恶劣的肿瘤代谢微环境中适应并介导免疫抑制,但其具体机制尚未明确。该研究通过空间多组学技术证实,肿瘤微环境中高浓度的氨是驱动 Treg 富集、抑制效应 T 细胞的核心代谢因子。Tregs 通过尿素循环解毒与精胺合成代谢两条特异性通路适应氨胁迫:一方面上调精氨酸琥珀酸裂解酶(ASL) 解除氨毒性;另一方面经FOXP3-SMS - 精胺 - PPARγ轴增强线粒体氧化磷酸化(OXPHOS)与免疫抑制功能。临床层面,抗 PD-1 治疗引发的肿瘤细胞死亡会通过转氨作用释放大量氨,进一步强化 Treg 并导致免疫治疗耐药。靶向氨代谢(如 GLUD1 抑制剂)可有效抑制 Treg、恢复抗肿瘤免疫,为下一代肿瘤免疫治疗提供全新策略。
Part.1
研究背景解读

Part.2
研究思路解析

空间定位:利用空间转录组 + 空间代谢组,定位肝癌中高谷氨酰胺分解、低尿素循环、高氨、高 Treg的共定位区域。 因果验证:体外氨处理、体内氨饮食 / 氨注射,证实氨选择性保护 Treg、杀伤效应 T 细胞。 机制拆解 解毒通路:氨→SRC3/STAT3→ASL 上调→尿素循环激活→解毒。 功能强化通路:氨→FOXP3→SMS 上调→精胺生成→结合 PPARγ→增强 OXPHOS 与抑制功能。 耐药机制:揭示抗 PD-1 诱导肿瘤死亡→转氨作用释放氨→强化 Treg→免疫耐药的新环路。 转化策略:验证GLUD1 抑制剂 + 抗 PD-1可阻断氨生成、克服耐药。
Part.3
主要研究结果展示

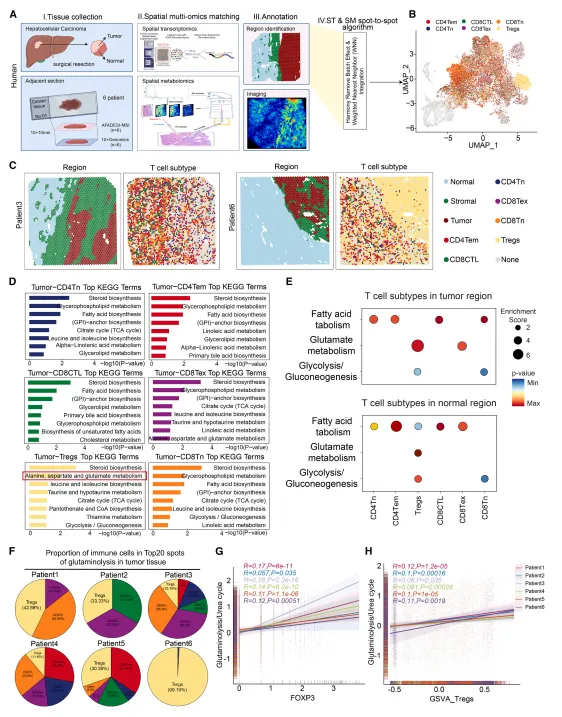
人类肝癌空间多组学显示:Treg 仅富集在谷氨酰胺分解活跃、尿素循环受抑的肿瘤区域,该区域氨浓度显著升高。 小鼠肝癌模型、肺癌 / 肠癌临床样本均验证:肿瘤氨浓度与 Treg 丰度呈强正相关。 体内注射氨、氨饮食均可直接促进肿瘤生长与 Treg 浸润。
低浓度氨(5 mM)不影响 Treg 存活,但显著抑制 CD8⁺/CD4⁺ T 细胞增殖并诱导凋亡。 Treg 在氨环境中维持胞内 pH、线粒体与溶酶体稳定,效应 T 细胞则出现严重损伤。 氨上调 Treg 的 FOXP3、PD-1、CTLA-4,直接增强其免疫抑制功能。
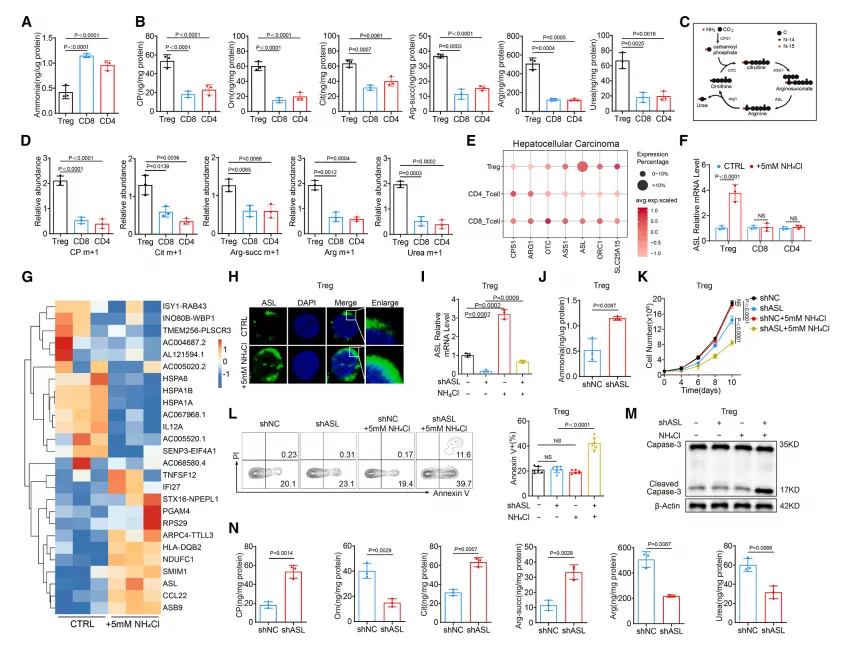
肿瘤浸润 Treg 特异性高表达ASL,外周 T 细胞无此特征。 氨通过SRC3-STAT3通路转录激活 ASL,启动尿素循环解毒。 敲低 / 敲除 ASL 后,Treg 丧失氨抗性,发生凋亡,肿瘤生长受抑。
氨通过 FOXP3 直接转录激活精胺合成酶(SMS),促进精胺生成。 精胺直接结合并激活 PPARγ,上调线粒体复合体基因,增强 OXPHOS。 X 射线晶体学证实:精胺与 PPARγ 的Glu319/Ser370/Glu371位点直接结合。 破坏该结合后,氨无法增强 Treg 的线粒体功能与抑制能力。
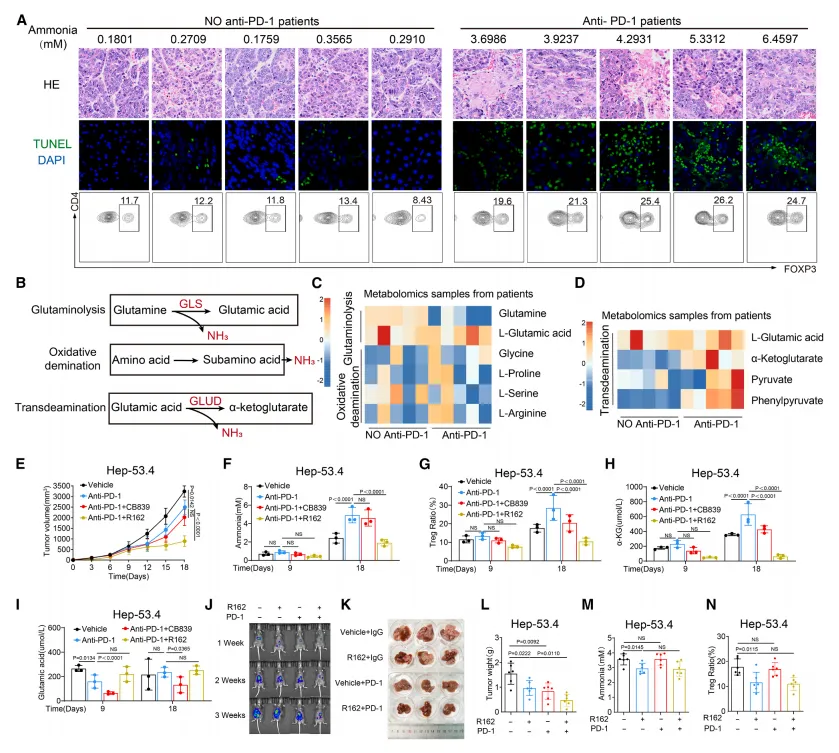
抗 PD-1 引发肿瘤细胞死亡,通过转氨作用(非谷氨酰胺分解)大量释放氨。 GLS1 抑制剂(CB839)无法降低该来源氨,GLUD1 抑制剂(R162)可显著阻断氨生成。 GLUD1 抑制剂 + 抗 PD-1联用:降低氨水平、减少 Treg、恢复 CD8⁺ T 细胞功能、克服耐药,在肝癌、肺癌、肠癌模型中均有效。
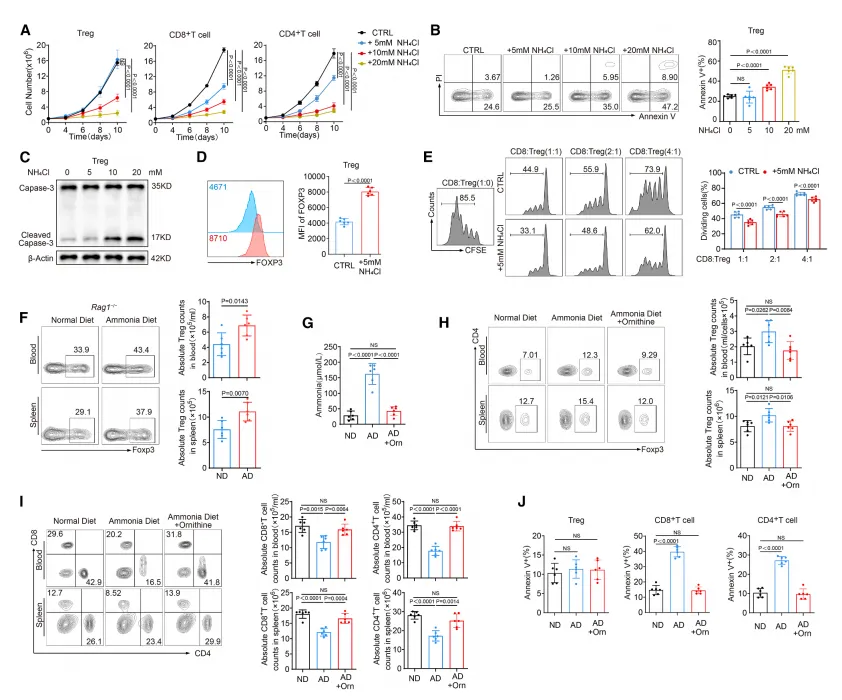
该研究通过体外细胞实验与体内动物模型验证,发现氨暴露可显著诱导 CD8⁺和 CD4⁺效应 T 细胞发生凋亡,而 Treg 细胞则表现出对氨诱导凋亡的显著抵抗性;这种抵抗性并未削弱 Treg 的 Foxp3 表达与免疫抑制功能,在含氨饮食的小鼠体内模型中,Treg 的比例与数量也能稳定维持,而效应 T 细胞则明显减少,提示氨的存在会选择性富集 Treg 细胞,进而加剧肿瘤微环境的免疫抑制状态。
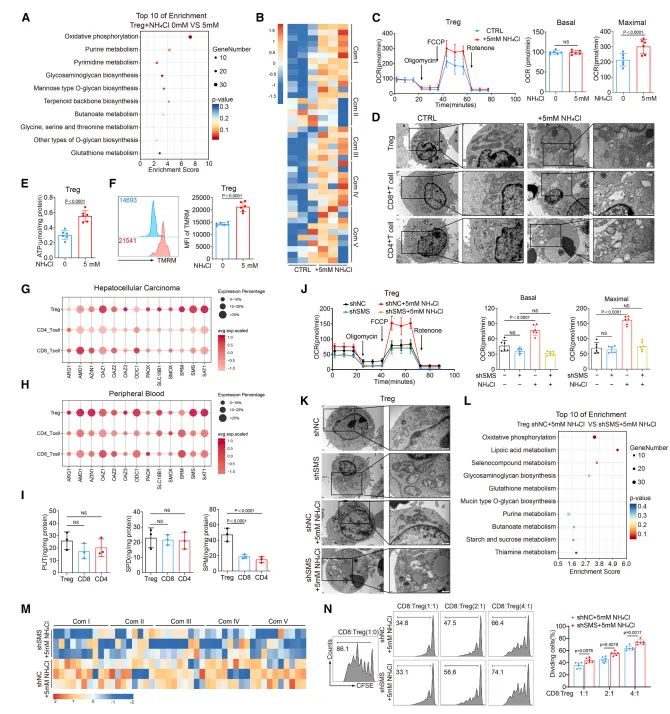
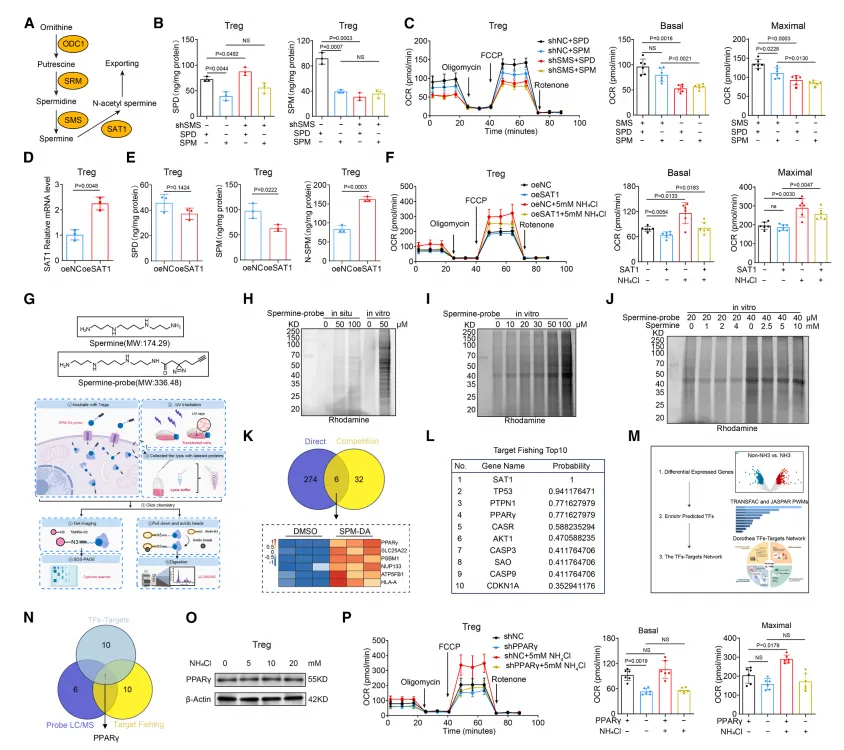
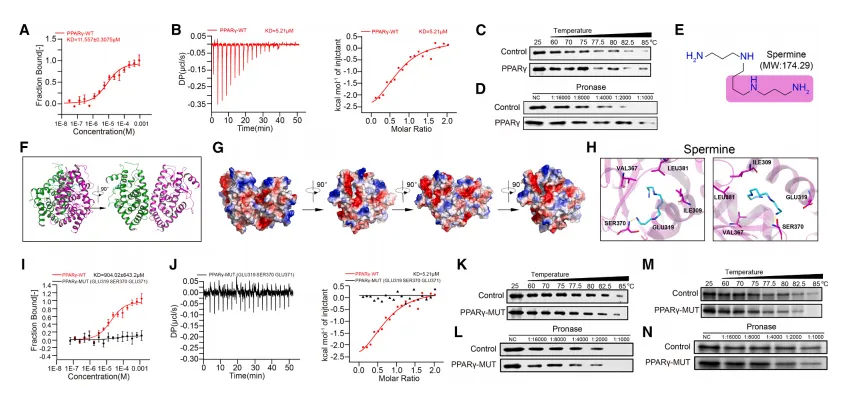
该研究通过多种生物物理实验(MST、ITC、CETSA、DARTS)证实,精胺可直接与 PPARγ 蛋白的配体结合口袋结合,显著增强其热稳定性与蛋白酶抗性;关键位点突变(E319A/S370A/E371A)会完全阻断精胺与 PPARγ 的相互作用,说明精胺通过直接结合并稳定 PPARγ 蛋白,进而调控 Treg 的线粒体复合物形成与氧化磷酸化功能。
Part.4
总 结

解毒轴:氨→SRC3/STAT3→ASL→尿素循环→存活。 功能轴:氨→FOXP3→SMS→精胺→PPARγ→OXPHOS→强免疫抑制。
随机文章
-
 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
- 南京银行北京分行
- 江苏懵了,扬州惊了,南京怎么就成了全国瞩目焦点?
- 南京市雨花台区母亲节订花送花微信:1340765130南京市雨花台区520情人节鲜花速递鲜花配送|站在我身后护我
- 2026.4.20-2026.4.24南京摩托车额度周参考均价
- 南京城市新版图展开!河西新城和江北新区为何不断放大价值
- 南京银行2025年每股分红0.529元;要21.93年回本.巴菲特:如果市场波动很大,市场是可以被利用的,而不是指导你将要发生什么.
- 【对话·校园媒体篇】南京大学卢璇:表达之外,她在时间里写信
- 南京东路淮海路黄金回收,魔都高价回收渠道
- IF8.3!6天接收2天见刊,南京中医药大学“国自然”热点联合“网药+分子对接”轻松发一区!
- 悲祭南京灵谷寺(南京行10)
