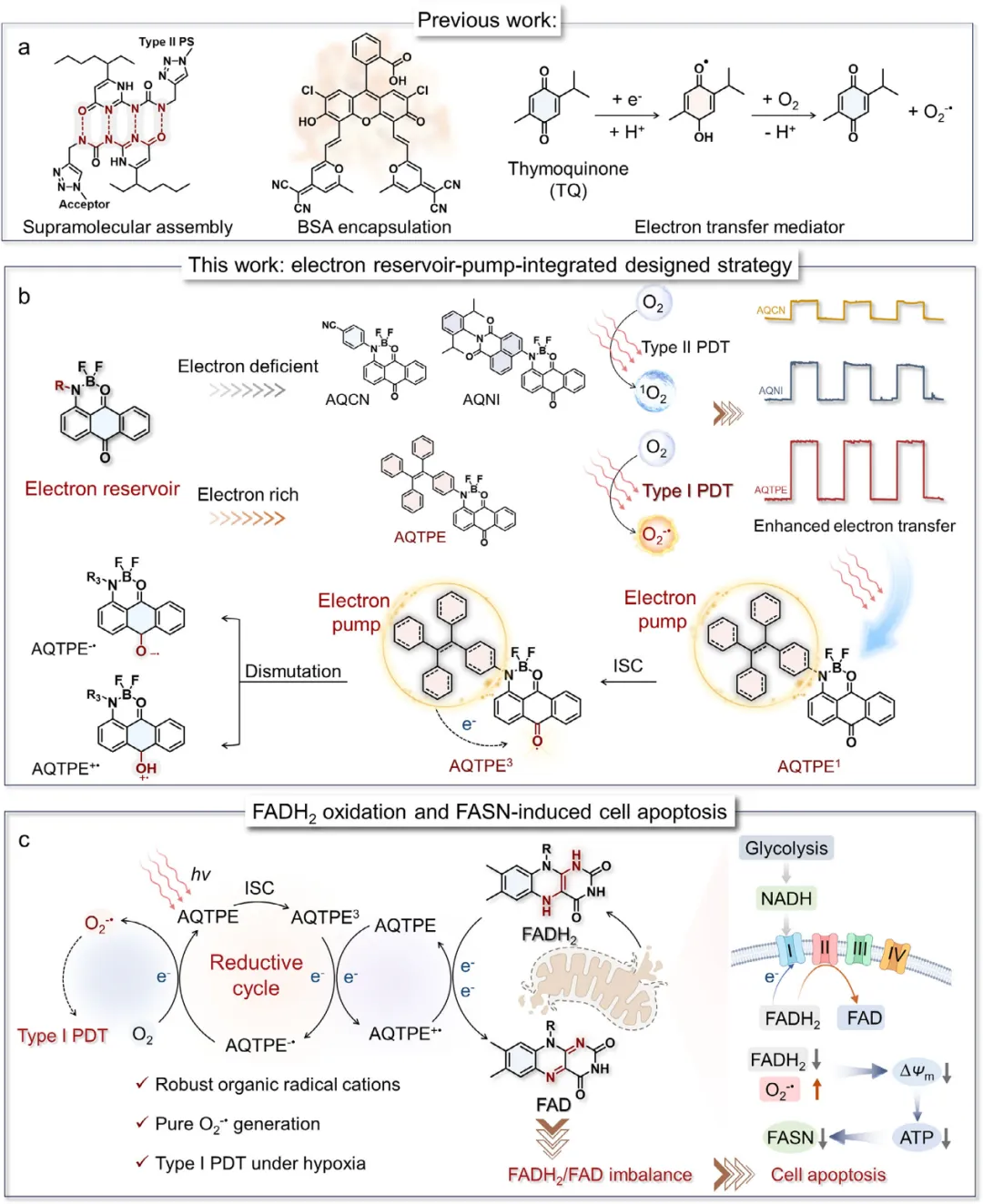
光动力疗法(PDT)具有卓越的选择性、非侵入性和低全身副作用,已成为一种前景广阔的肿瘤治疗策略。PDT的治疗机制主要依赖于光敏剂在光照下产生细胞毒性活性氧(ROS),该过程通过两种主要途径实现。在传统的II型途径中,光敏剂激发态三重态(Tn)的能量转移至分子氧(O₂),生成单线态氧(¹O₂);这一过程高度依赖O₂,且是大多数传统光敏剂的主要作用方式。相比之下,I型途径涉及光诱导的电子转移,可产生超氧阴离子(O₂⁻•)和羟基自由基(•OH),其对O₂的依赖性显著较低。由于I型光敏剂具有优异的低氧耐受性,因此尤其适用于对抗实体瘤特有的缺氧微环境。
尽管有此潜力,开发具有内在强电子转移能力的光敏剂仍然是一项根本性挑战。目前的策略,例如生物素化、阳离子化、重原子取代或对II型光敏剂进行结构修饰以诱导I型反应性,仅取得了有限的成功。最近,人们探索了超分子共组装方法,通过将II型光敏剂与电子中介体配对来提高电子转移效率(Scheme 1a)。例如,将百里醌(TQ)与传统光敏剂共同封装,可促进电子穿梭,从而在低氧条件下将ROS的产生从II型途径转变为I型途径。尽管这些多组分系统代表了概念上的进步,但由于给体与受体单元之间的空间分离导致分子间电子转移效率低下,其性能常常受到影响。
蒽醌(AQ)衍生物因其缺电子的羰基骨架,可在光激发下稳定自由基中间体,已被广泛用作光动力疗法(PDT)中的氧化还原活性发色团。包括天然产物如大黄素在内的经典代表已被报道可作为光敏剂;然而,它们主要通过能量转移主导或II型/I型混合途径发挥作用,其活性氧(ROS)的产生在很大程度上仍然依赖于氧气和被动的激发态过程。尽管这些研究强调了蒽醌核心的电子接受特性,但其光动力性能受限于缓慢且受扩散控制的电子转移,导致在低氧条件下自由基生成不足。更为关键的是,现有基于蒽醌的光敏剂被设计为孤立的电子受体,而非集成的氧化还原系统,缺乏定向电荷分离的内在驱动力。为了克服这些限制并充分释放蒽醌骨架纯I型光动力的潜力,作者从醌类氧化还原介体TQ(百里醌)中汲取了灵感。作者假设:将蒽醌作为一个固有的电子储库,与一个互补的给电子基团整合,将有望构建出一种单组分光敏剂,能够实现高效的分子内电子转移和不依赖氧气的自由基生成。
在此,作者报道了一种单分子电子储库-泵光敏剂AQTPE,该光敏剂在低氧条件下实现了高效的I型光动力活性。在该设计中,蒽醌(AQ)核心作为电子储库以稳定自由基阴离子,而富电子的四苯基乙烯(TPE)单元则作为电子泵促进空穴抽取,从而为光诱导歧化反应和自由基离子对的形成建立了内在驱动力(图1b)。由此产生的阳离子自由基可直接氧化关键的代谢辅酶(如黄素腺嘌呤二核苷酸,FADH₂),而阴离子自由基则将O₂还原为O₂⁻•,从而实现无需外部电子供体的双重活性氧(ROS)生成机制。这种双重反应性不仅放大了细胞毒性ROS的产生,还建立了一种代谢靶向的光动力疗法策略。FADH₂的氧化会导致脂肪酸合成酶(FASN)受到抑制,从而破坏快速增殖癌细胞所必需的脂质生物合成;同时,它还会损害线粒体电子传递链的功能,进而引发线粒体功能障碍和细胞凋亡(图1c)。此外,转录组分析揭示,AQTPE介导的光激活会失调氧化应激反应、NADH氧化以及Bcl介导的凋亡通路。据作者所知,这是首次在I型光动力疗法中证明FASN抑制作为一种治疗机制,为将光化学与代谢干预相整合提供了新的分子蓝图。作者的工作建立了一种基于外围电负性调控的通用设计原则,用于构建单组分电子储库-泵光敏剂,为下一代具有强临床转化潜力的低氧适应性I型光动力疗法铺平了道路。
方案1 (a) 已报道的I型光敏剂设计策略。(b) 图示说明蒽醌(AQ)核心作为“电子泵”、四苯基乙烯(TPE)单元作为“电子储库”,通过增强电子转移生成自由基离子对。(c) 图示说明光诱导歧化反应用于黄素腺嘌呤二核苷酸(FADH₂)氧化及超氧阴离子(O₂⁻•)生成。
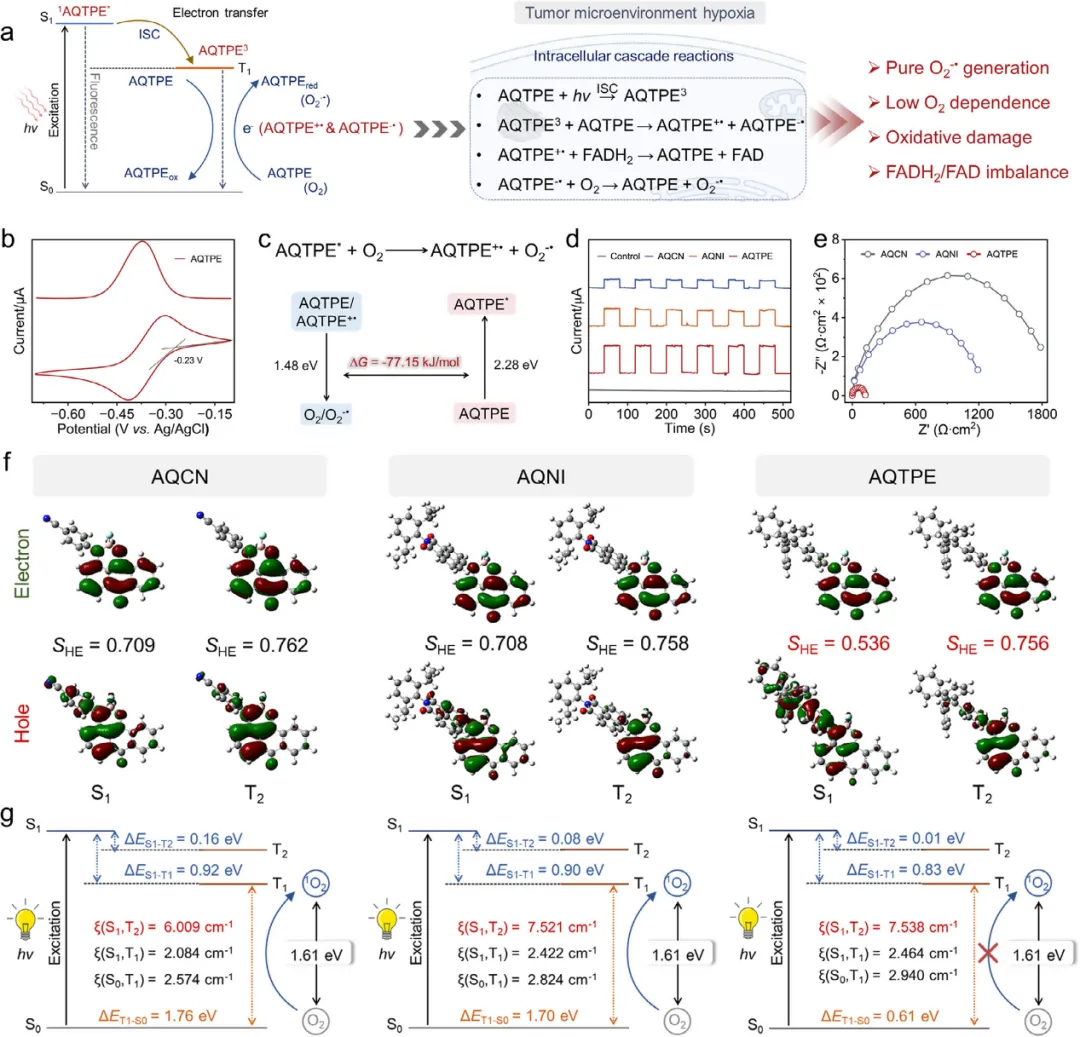
图1 (a) AQCN纳米颗粒(NPs)、AQNI NPs和AQTPE NPs的紫外-可见吸收光谱。(b) 三种NPs在PBS中的流体动力学尺寸。插图为三种NPs的扫描电镜(SEM)图像。(c) 三种NPs的Zeta电位。(d) 结晶紫、孟加拉红(RB)、AQCN、AQNI和AQTPE的ROS产量及能隙比较的雷达图。(e) 结晶紫NPs、RB NPs、AQCN NPs、AQNI NPs和AQTPE NPs的总ROS生成量,(f) 单线态氧(¹O₂)生成量,以及(g) 超氧阴离子(O₂⁻•)生成量。(h) TEMP和(i) DMPO在不同条件下的电子自旋共振(ESR)谱图。(j) 260 nm处吸光度增加和(k) 339 nm处吸光度减少所指示的NADH光氧化时间进程(NADH: 100 µM)。(l) 450 nm处吸光度减少所指示的FADH₂光氧化时间进程(FADH₂: 50 µM)。(m) 光催化FADH₂/FAD转化及其特征吸收峰。(n) 基于密度泛函理论(DFT)计算得到的AQCN、AQNI和AQTPE在基态(S₀)下的最高占据分子轨道(HOMO)和最低未占分子轨道(LUMO)。
图2 (a) AQTPE NPs光激发I型光动力疗法过程示意图。(b) AQTPE在干燥二氯甲烷(DCM)中的循环伏安(CV)曲线。(c) AQTPE的吉布斯自由能(∆G)变化。(d) 在0.10 V偏压、白光照射下,AQCN、AQNI和AQTPE的光电流响应谱。(e) 光照下AQCN、AQNI和AQTPE的电荷转移电阻。(f) AQCN、AQNI和AQTPE的前沿激发单重态和三重态的自然跃迁轨道。(g) 基于含时密度泛函理论(TD-DFT)计算得到的AQCN、AQNI和AQTPE的前沿分子轨道、单重态-三重态能隙及系间窜越(ISC)形成机制示意图。
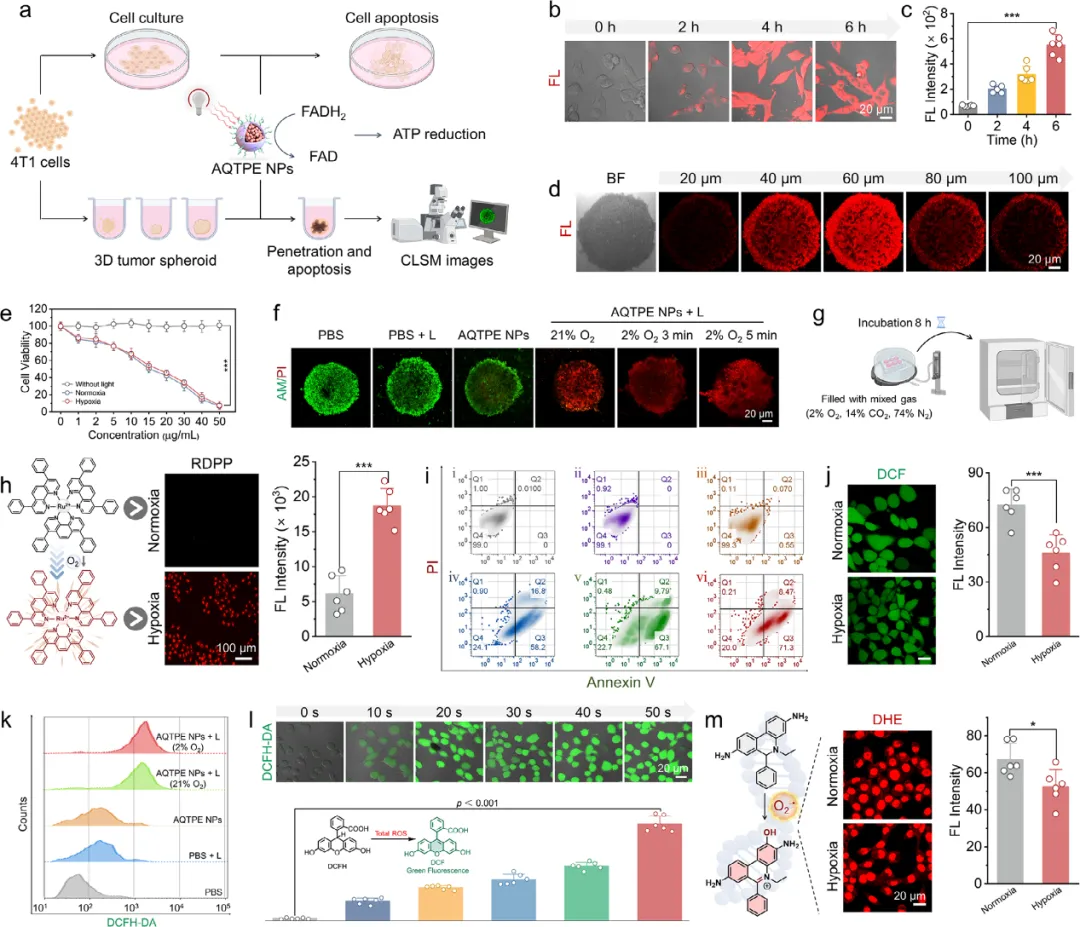
图3 (a) AQTPE NPs诱导癌细胞凋亡及3D肿瘤球形成模型示意图。(b) 4T1细胞对AQTPE NPs的摄取情况(激发波长λex = 533 nm,发射波长λem = 660–800 nm)。(c) AQTPE NPs在细胞摄取过程中荧光强度的定量分析。(d) AQTPE NPs穿透4T1肿瘤球的代表性z轴层扫共聚焦图像。(e) 不同浓度AQTPE NPs在常氧和低氧条件下、有无光照处理的MTT实验结果。(f) 不同处理后4T1三维多细胞球中活/死细胞的共聚焦成像。Calcein-AM产生的绿色荧光代表活细胞,PI产生的红色荧光代表死细胞。(g) 细胞内低氧微环境模型示意图。(h) 使用低氧探针RDPP指示的细胞内O₂浓度。左侧为检测原理示意图。(i) 流式细胞术分析4T1细胞凋亡情况。细胞与AQTPE NPs孵育后进行光照处理:i) PBS,ii) PBS + 光照,iii) AQTPE NPs,iv) AQTPE NPs + 光照(30 µg mL⁻¹),v) 常氧条件下AQTPE NPs + 光照(50 µg mL⁻¹),vi) 低氧条件下AQTPE NPs + 光照(50 µg mL⁻¹)。(j) 使用DCFH-DA作为总ROS指示剂检测4T1细胞内总ROS水平。(k) 流式细胞术检测不同处理后4T1细胞内的ROS水平。(l) 4T1细胞与AQTPE NPs孵育并在白光(80 mW cm⁻²,50秒)照射后,DCFH-DA染色的时间依赖性图像。不同照射时间下的荧光强度见下图。(m) 常氧和低氧条件下,经AQTPE NPs + 光照处理后,使用DHE作为特异性探针进行的细胞内O₂⁻•成像。左侧为检测原理示意图。误差条表示平均值±标准差(n = 6),**p < 0.001,p < 0.05。
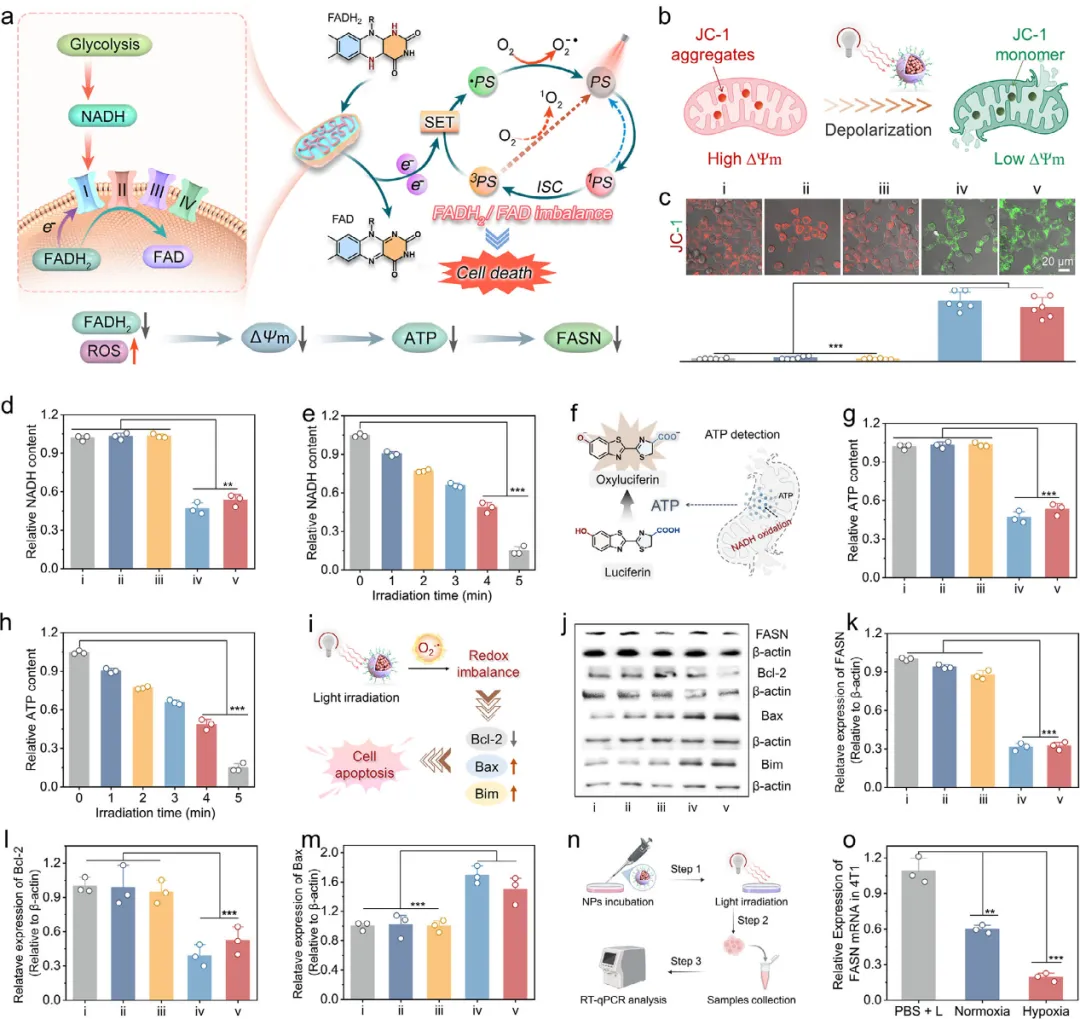
图4 (a) 示意图展示AQTPE NPs诱导线粒体电子传递链的破坏。(b) JC-1检测原理示意图。(c) 不同处理后4T1癌细胞的JC-1染色。光照剂量:80 mW cm?2,8分钟。绿色荧光代表JC-1单体(低线粒体膜电位,low MMP),红色荧光代表JC-1聚集体(高线粒体膜电位,high MMP)。下方为对应的红/绿荧光比值的半定量分析。误差条表示平均值±标准差(n=6),*p < 0.001。(d) 不同处理后细胞内NADH的消耗。(e) AQTPE NPs处理后并光照不同时间(80 mW cm?2)的4T1细胞中NADH水平。(f) ATP检测方法示意图。(g) 不同处理后4T1细胞中的ATP水平。(h) 不同光照时间下AQTPE NPs处理后的4T1细胞中ATP含量。(i) 所提出的光催化反应及其导致的FADH?/FAD失衡和ROS生成示意图,这些作用破坏线粒体电子传递链(ETC)并诱导细胞凋亡。(j) 不同处理后4T1细胞中FASN、Bcl-2、Bax和Bim蛋白表达的蛋白质印迹(WB)分析。基于蛋白质印迹结果的(k) FASN、(l) Bcl-2和(m) Bax蛋白表达的相应半定量分析。(i) PBS,(ii) PBS + L,(iii) AQTPE NPs,(iv) 常氧条件下AQTPE NPs + L,(v) 低氧条件下AQTPE NPs + L。条带强度通过Image J分析。数据表示为三次独立实验的平均值和标准差(SD)。(n) AQTPE NPs体外光动力激活的一般流程。(o) 常氧或低氧条件下,经AQTPE NPs + L处理的4T1细胞中FASN mRNA表达水平。误差条表示平均值±标准差(n=3),*p < 0.001,**p < 0.01。
图5 (a) 热图总结了RNA-seq样本之间的皮尔逊相关系数:4T1细胞与PBS共孵育并光照5分钟(C-1、C-2和C-3);4T1细胞与AQTPE NPs共孵育并光照5分钟(1-1、1-2和1-3)。(b) 与PBS+光照组相比,AQTPE NPs处理后经光照的4T1细胞中上调(红色)和下调(蓝色)基因的火山图(|log2倍数变化| ≥ 1,p < 0.05)。(c) 聚类分析和热图展示了AQTPE NPs+光照处理组与PBS+光照处理组4T1细胞中差异表达基因的总体情况(n = 3,p ≤ 0.05)。每一行代表一个样本,每一列代表一个基因。颜色表示基因的表达水平。(d) 与PBS+光照组相比,AQTPE NPs处理后经光照的4T1细胞中上调基因的KEGG富集分析和(e) 基因本体论(GO)富集分析(FDR < 0.05)。气泡的大小代表每个条目中的基因数量。(f) AQTPE NPs介导的PDT在4T1细胞中诱导的前三个上调通路的GSEA分析(FDR < 0.05)。NES,归一化富集分数。
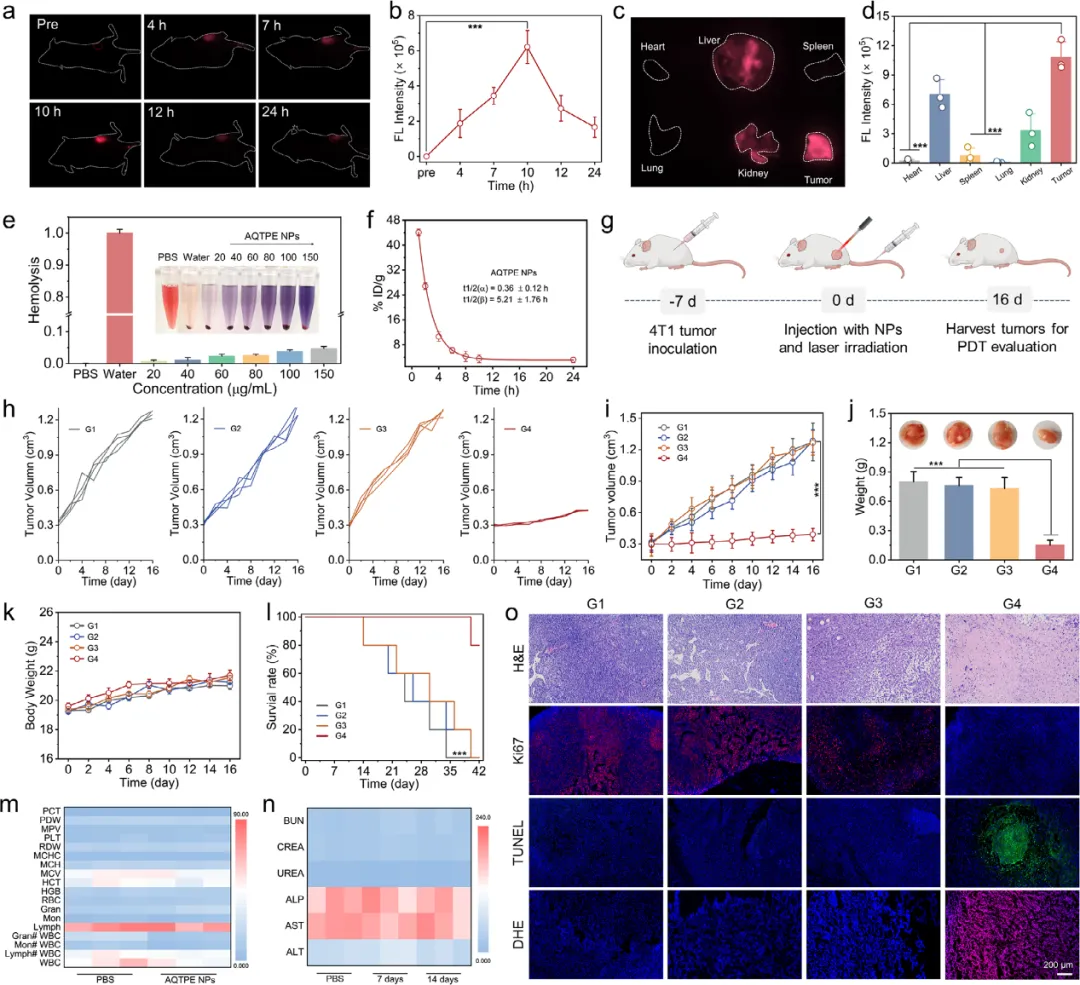
图6 (a) 不同时间点AQTPE NPs的体内荧光成像。(b) 反映AQTPE NPs体内循环和肿瘤蓄积的时间依赖性荧光强度变化的定量分析。(c) AQTPE NPs在肿瘤和主要器官中分布的离体荧光成像。(d) 比较主要器官和肿瘤组织中AQTPE NPs蓄积量的荧光强度定量分析。误差条表示平均值±标准差(n=3),*p < 0.001。(e) 不同浓度AQTPE NPs的溶血率。(f) AQTPE NPs的药代动力学曲线。(g) 肿瘤接种、AQTPE NPs注射、光照及肿瘤生长监测的示意图。(h) 不同处理后各组小鼠的个体肿瘤体积。(i) 16天内不同处理后各组小鼠的肿瘤体积变化。(j) 不同处理后各组小鼠的生存率。(k) 不同处理后各组小鼠的相对体重变化。(l) 不同处理结束时的平均肿瘤重量(平均值±标准差,n=4)。(m) 不同处理组(PBS组和AQTPE NPs组)健康小鼠的血液生化参数和(n) 血常规参数。(o) 不同处理后肿瘤组织的H&E染色、Ki67、TUNEL和DHE染色。G1,PBS组;G2,PBS+光照组;G3,AQTPE NPs组;G4,AQTPE NPs+光照组。误差条表示平均值±标准差(n=4),*p < 0.001。
总之,作者开发了一种外围电负性调控策略,用于构建一系列蒽醌衍生光敏剂,该策略能够精确控制分子内电子转移,并即使在严重低氧条件下也能实现高效的I型光动力疗法。通过将富电子的四苯基乙烯(TPE)单元引入蒽醌骨架,作者建立了一个分子内电子储库-泵系统,显著增强了电子转移并放大了超氧阴离子(O₂⁻•)的生成。机理研究表明,光激发的AQTPE经历了高效的S₁→Tₙ系间窜越(ISC)过程,随后发生双自由基离子对级联反应:阴离子自由基将分子氧还原为O₂⁻•,而阳离子自由基则氧化代谢辅酶FADH₂。这种双氧化还原反应活性破坏了细胞内氧化还原平衡,抑制了脂肪酸合成酶(FASN)的表达,并诱导线粒体功能障碍,最终导致代谢靶向的细胞凋亡。理论计算进一步表明,AQTPE具有极小的ΔEST(单重态-三重态能隙)和较大的自旋轨道耦合(SOC)常数,这一特征不利于传统的能量转移,但有利于ISC和自由基离子的形成,从而确保了高度选择性的I型途径。体外和体内评估均证实,AQTPE纳米颗粒(NPs)在低氧条件下表现出强大的光动力细胞毒性和显著的肿瘤抑制效果。总体而言,本研究不仅引入了一种低氧适应性光敏剂,还提出了一种通用的分子设计原则——外围电负性调控与电子储库-泵工程相结合——用于下一代I型光动力疗法。通过将光化学自由基生成与代谢脆弱性直接关联,作者的工作为开发具有临床转化潜力的代谢靶向光敏剂提供了新的见解。
文献链接:https://doi.org/10.1002/anie.9985126
点击“阅读原文”直达上述文章











 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
