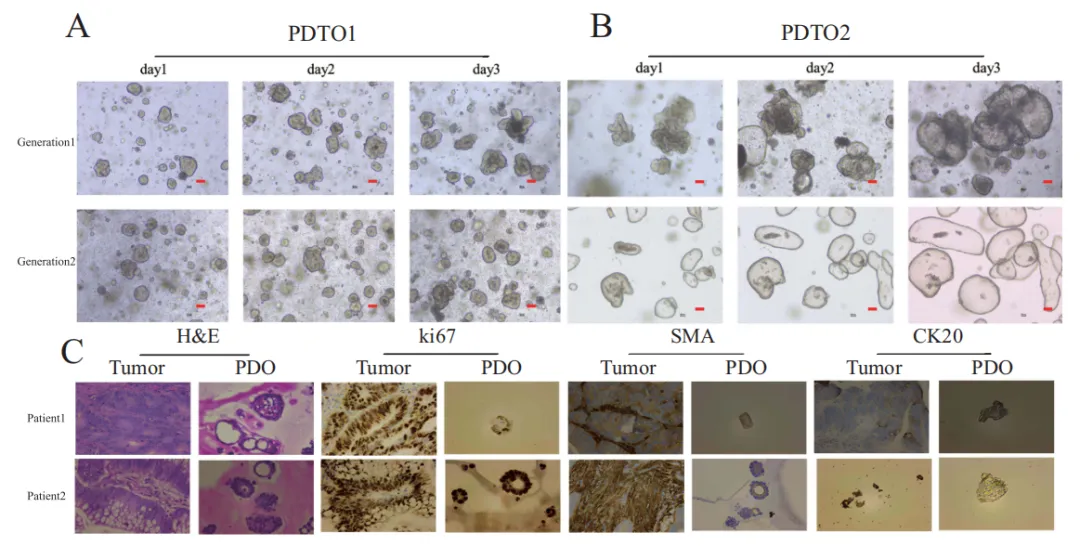
研究首先成功构建 24 例结直肠癌患者来源类器官(PDTOs),形态包括囊状、致密球状等,可稳定传代(Fig.1A、B)。HE 染色与免疫组化显示,类器官与原发肿瘤结构高度一致,Ki67、CK20 强阳性,SMA 阴性,完美保留肿瘤特征(Fig.1C)。全外显子测序进一步证实,类器官与对应肿瘤在SNP、InDel、CNV上高度匹配,基因特征一致(Fig.2),确保后续药效预测真实可靠。
Fig.1 完成结直肠癌 PDTO 模型的建立与表征,为药物筛选提供可靠平台。A–B 动态观察显示,原代及传代类器官呈囊状、致密球状等典型形态,随培养时间体积增大、结构稳定,不同患者来源类器官具备形态异质性,符合临床肿瘤多样性特征。C 对两类器官与对应原发肿瘤进行组织学与标志物对比,HE 染色显示类器官高度复刻肿瘤组织的腺体结构与核异型性;IHC 结果证实,类器官与肿瘤组织均高表达增殖标志物 Ki67 及结直肠癌特异性标志物 CK20,间质标志物 SMA 呈阴性,排除成纤维细胞污染。该图从形态、组织学、分子水平全面验证 PDTO 准确保留患者肿瘤的生物学特征,可真实反映体内药物响应,为后续紫杉醇敏感性检测与代谢组关联分析奠定坚实模型基础。
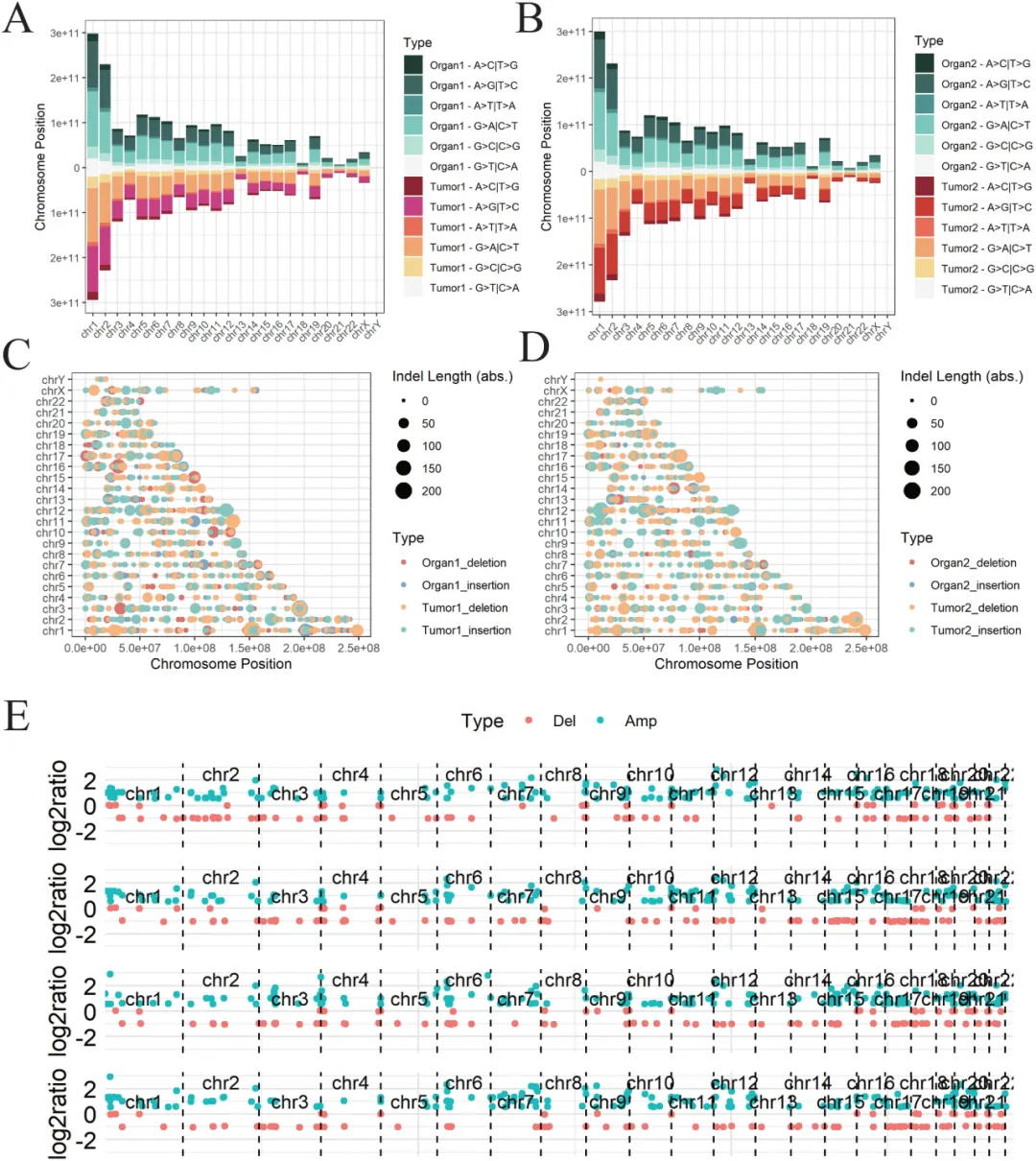
Fig.2 从全基因组层面证实 PDTO 完美复刻患者肿瘤突变特征。A–B 单核苷酸变异(SNV)分析显示,两类器官与其匹配肿瘤组织的突变类型、比例高度一致,无明显偏差。C–D 插入缺失(InDel)检测表明,类器官与肿瘤在 InDel 长度、分布及突变频率上高度重合,维持基因组结构稳定性。E 拷贝数变异(CNV)谱显示,类器官与对应肿瘤的扩增、缺失区域完全匹配,不同样本间存在个体化差异。全外显子测序结果证明,PDTO 在 SNV、InDel、CNV 三个维度均高度保留原发肿瘤的基因组特征,无培养诱导的额外畸变,确保药物敏感性差异源于患者本身遗传与代谢背景,而非体外驯化,为 “类器官结果→临床意义” 的转化提供强力遗传学支撑。
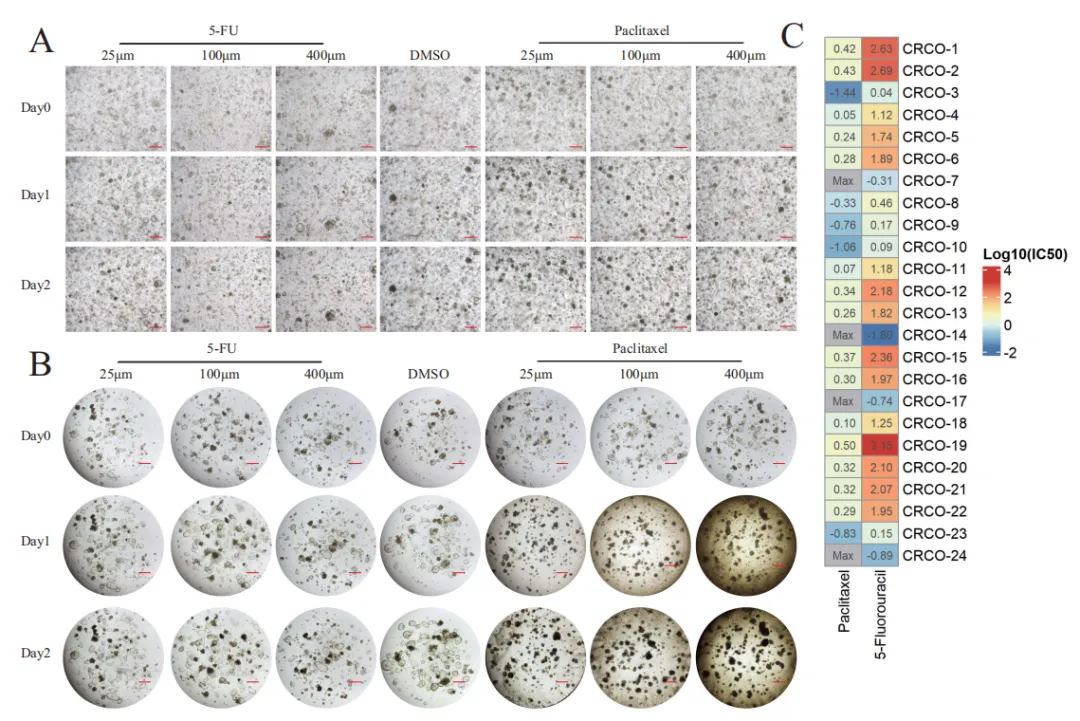
在 24 例类器官中开展紫杉醇与 5‑FU 对比测试,结果显示多数类器官对紫杉醇更敏感,但个体差异极大(Fig.3A、B)。研究统计 log10 (IC50) 并绘制热图,清晰呈现从高度敏感到完全耐药的连续分布(Fig.3C)。亚组分析显示,年龄、性别、BMI、分期均不能解释这种差异,提示疗效差异来自代谢等内在因素。
Fig.3 在24 例结直肠癌 PDTO中系统比较紫杉醇与 5‑FU 的药效差异。A–B 高内涵成像显示,5‑FU 处理后类器官形态、密度无明显改变,生长抑制微弱;而紫杉醇组细胞明显黑化、边界模糊、结构崩解,呈现显著杀伤。C 热图展示 24 例 PDTO 对两种药物的 Log10 (IC50),多数样本对紫杉醇更敏感,IC50 跨度大,提示个体差异显著;部分样本 IC50 超出检测上限,归为截尾值(Max)。亚组分析表明,BMI、年龄、性别、肿瘤分期均与紫杉醇敏感性无显著关联,排除临床混杂因素干扰。该图首次在大样本 PDTO 中证明,紫杉醇对难治性结直肠癌具有潜在治疗价值,且响应差异不由临床病理特征决定,提示代谢等内在因素是关键调控因子,引导研究转向代谢组筛选。
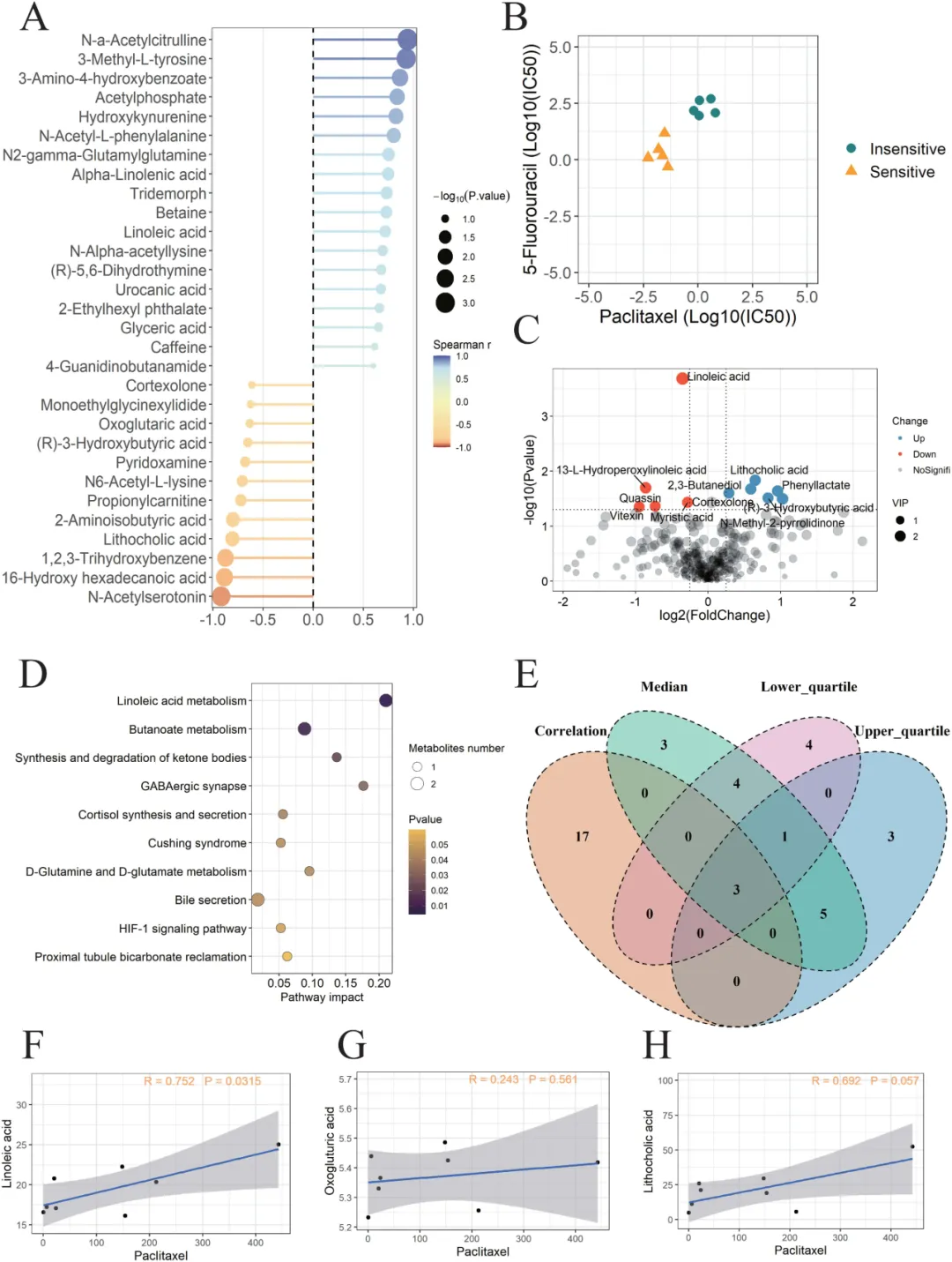
研究选取 18 对有效数据,分为发现组与验证组,进行血清非靶向与靶向代谢组分析(Fig.4)。结果显示,亚油酸与紫杉醇 IC50 呈显著正相关,水平越高,耐药越强。KEGG 富集提示亚油酸代谢通路是关键通路,且在验证队列中相关性稳定可重复(Fig.4F–H)。
Fig.4 联合PDTO 药效与血清代谢组,鉴定出调控紫杉醇敏感性的核心代谢物。A 气泡图展示与 IC50 相关性最强的 30 种代谢物,亚油酸(LA)相关性最显著。B 散点图直观呈现 PDTO 对紫杉醇与 5‑FU 响应的异质性。C 火山图显示敏感组与耐药组的差异代谢物分布。D KEGG 富集表明,亚油酸代谢是最显著富集通路。E 多策略交叉分析锁定 LA、酮戊二酸、石胆酸 3 个候选物。F–H 靶向定量验证,仅 LA 在验证队列中与 IC50 呈稳定正相关(R=0.752),高 LA 对应高 IC50(耐药)。逐一剔除样本的敏感性分析证实,LA 关联不受单一样本驱动。该图严谨证明血清 LA 是预测结直肠癌紫杉醇耐药的代谢标志物,为机制探索提供明确靶点。
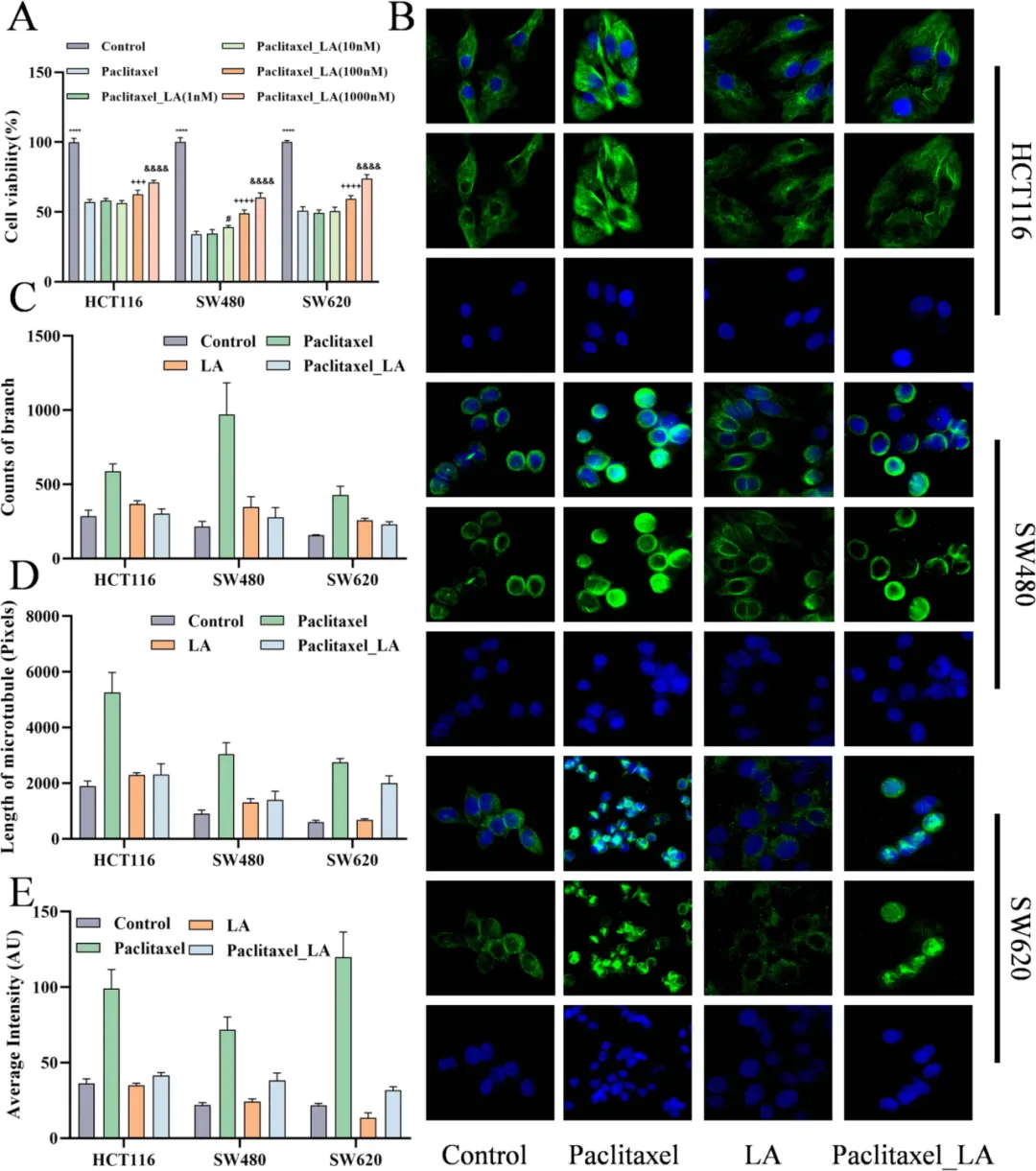
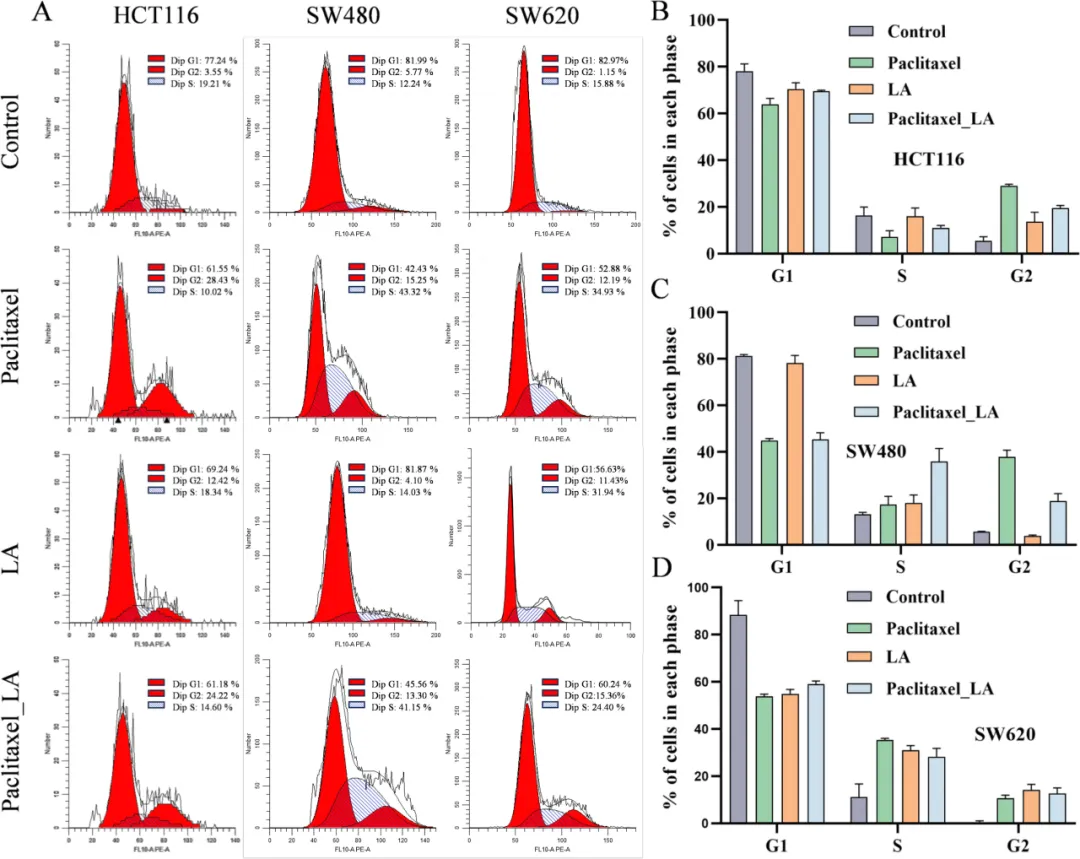
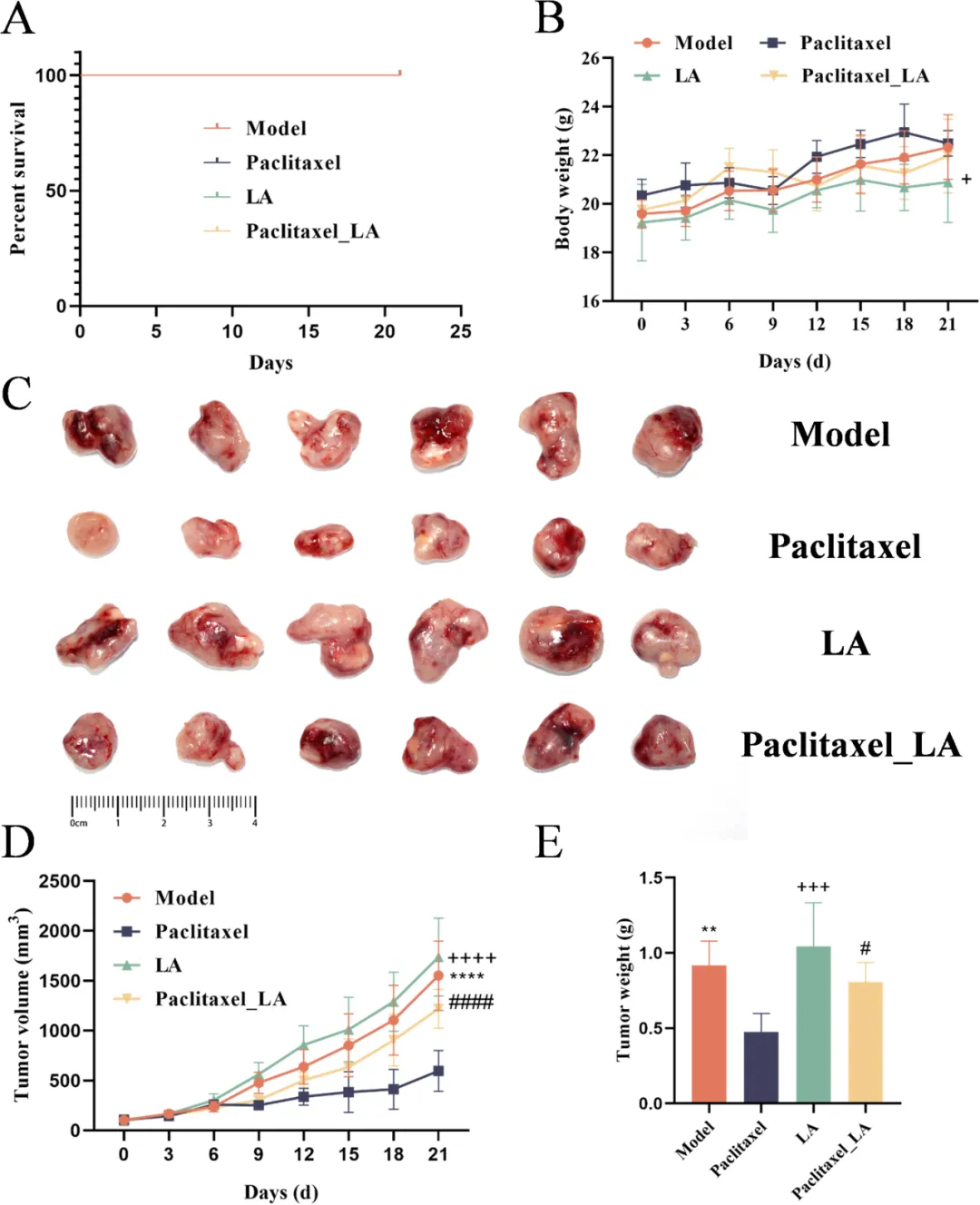
细胞实验显示,亚油酸可剂量依赖性逆转紫杉醇的杀伤作用(Fig.5A),并干扰微管聚集、降低稳定化程度(Fig.5B–E),减弱 G2/M 周期阻滞(Fig.6)。小鼠荷瘤模型进一步证实,联合亚油酸会显著降低紫杉醇抑瘤率,肿瘤体积与重量明显回升(Fig.7),完整证明其促耐药作用。
Fig.5 在细胞水平解析 LA 拮抗紫杉醇的分子机制。A CCK‑8 结果显示,LA 呈浓度依赖性逆转紫杉醇对 HCT116、SW480、SW620 的杀伤,100 nM 即可显著恢复细胞活力。B 免疫荧光显示,紫杉醇诱导微管高度聚集、致密化;LA 单独处理微管结构松散;联合组微管聚合程度显著低于紫杉醇单药,证明 LA 削弱紫杉醇对微管的稳定作用。C–E 定量分析证实,LA 显著降低紫杉醇诱导的微管分支数、总长度与荧光强度。结果表明,紫杉醇通过稳定微管发挥抗肿瘤作用,而 LA 通过改变微管动力学、抑制过度聚合,直接拮抗紫杉醇核心机制,从细胞骨架层面揭示 LA 介导耐药的结构基础,为后续细胞周期验证提供方向。
Fig.6 从细胞周期层面进一步验证 LA 的耐药效应。A 流式结果显示,紫杉醇可显著诱导 HCT116、SW480 细胞 G2/M 期阻滞,SW620 细胞 S/G2 期滞留,触发有丝分裂停滞。B–D 定量分析表明,LA 单独对周期影响微弱;但联合处理时,LA 显著降低 G2/M 比例,使细胞重新进入周期,缓解紫杉醇造成的分裂阻断。该结果与微管实验完全呼应:LA 破坏紫杉醇对微管的稳定作用,使纺锤体检查点无法持续激活,细胞绕过阻滞继续分裂,从而耐受紫杉醇杀伤。该图完整证明,LA 通过干扰微管→解除周期阻滞的通路,介导结直肠癌对紫杉醇的耐药,完成体外机制闭环。
Fig.7 在CT26 小鼠移植瘤模型中完成体内验证,为临床转化提供关键证据。A 生存曲线显示各组均无死亡,实验安全。B 体重监测表明治疗无明显毒性,耐受性良好。C–E 肿瘤图像、生长曲线与瘤重结果一致:紫杉醇单药显著抑制肿瘤生长;LA 单药无抑癌效果;紫杉醇 + LA 组肿瘤体积与重量显著高于紫杉醇单药组,抑瘤效果大幅下降。线性混合效应模型证实,治疗与时间存在显著交互作用,LA 持续拮抗紫杉醇疗效。该图在活体水平严格证明,体内高 LA 水平会直接降低紫杉醇的结直肠癌治疗效果,与 PDTO、细胞、机制结论完全统一,支持将血清 LA 作为紫杉醇用药前的分层标志物,指导临床精准用药。
这项研究以结直肠癌类器官为核心平台,首次建立 “血清亚油酸 — 紫杉醇疗效” 的预测关系,为临床提供简单、低成本的疗效预判指标。未来可通过检测患者血清亚油酸水平,提前筛选适合紫杉醇治疗的优势人群,避免无效化疗与过度治疗。
同时,研究揭示脂质代谢影响微管稳态的新机制,提示控制亚油酸水平、干预脂代谢可能成为逆转紫杉醇耐药的新策略。类器官 + 代谢组的组合模式,也可推广至奥沙利铂、伊立替康等其他药物,快速挖掘更多疗效标志物,推动结直肠癌进入 “代谢分型 + 精准用药” 的新时代,让每一位患者都能用上最适合的化疗方案。