2026 年 2 月 16 日,来自南京工业大学的Jianjian Tong等人在Journal of Agricultural and Food Chemistry上发表了一篇题为Designing a Sucrose Phosphorylase from Leuconostoc mesenteroides for the Highly Selective Production of EGCG-4′‑O‑α‑D‑Glucopyranoside的研究性论文。主要的绿茶多酚表没食子儿茶素没食子酸酯具有有益的抗氧化和抗炎活性,但其溶解度和稳定性较差。其单糖基化衍生物(−)-表没食子儿茶素没食子酸酯4′-O-α-D-吡喃葡萄糖苷部分克服了这些限制。在本研究中,工程化了肠膜明串珠菌蔗糖磷酸化酶以高效生产EGCG-G1。通过环工程、共识设计和Rosetta Dock设计创建的三重突变体M3(T219L/E393I/N335G),在30°C下表现出比野生型高4.09倍的转糖基活性,在45°C下半衰期延长1.69倍,并且区域选择性显著提高。在30°C、pH 6.0的补料分批反应中,25 g/L EGCG在24小时内转化,产生了31.11 g/L(产率91.92%)的EGCG-G1,纯度为87.27%。这种半理性设计策略增强了LmSPase的关键特性,并为EGCG-G1的生产提供了一种有效的生物催化剂。
Introduction
绿茶是一种广泛饮用的饮料,大量体外和体内研究表明,绿茶中的多酚具有促进健康的作用。绿茶中主要的多酚化合物是儿茶素,包括(−)-表没食子儿茶素没食子酸酯(EGCG)、(−)-表没食子儿茶素(EGC)、(−)-表儿茶素没食子酸酯(ECG)和(−)-表儿茶素(EC)。在这些儿茶素中,EGCG约占儿茶素总含量的40%。EGCG分子结构中丰富的酚羟基使其与其他酚类化合物相比具有增强的特性,包括抗氧化活性、预防细菌感染、抗炎作用、降低血糖和血脂水平、以及神经保护作用。尽管天然EGCG具有相当大的实用价值,但其应用受到稳定性低、吸收差、和溶解度相对较低等挑战的限制。因此,可以通过化学或酶法修饰引入官能团,以产生更稳定且易于吸收的衍生物,从而增强其特性。
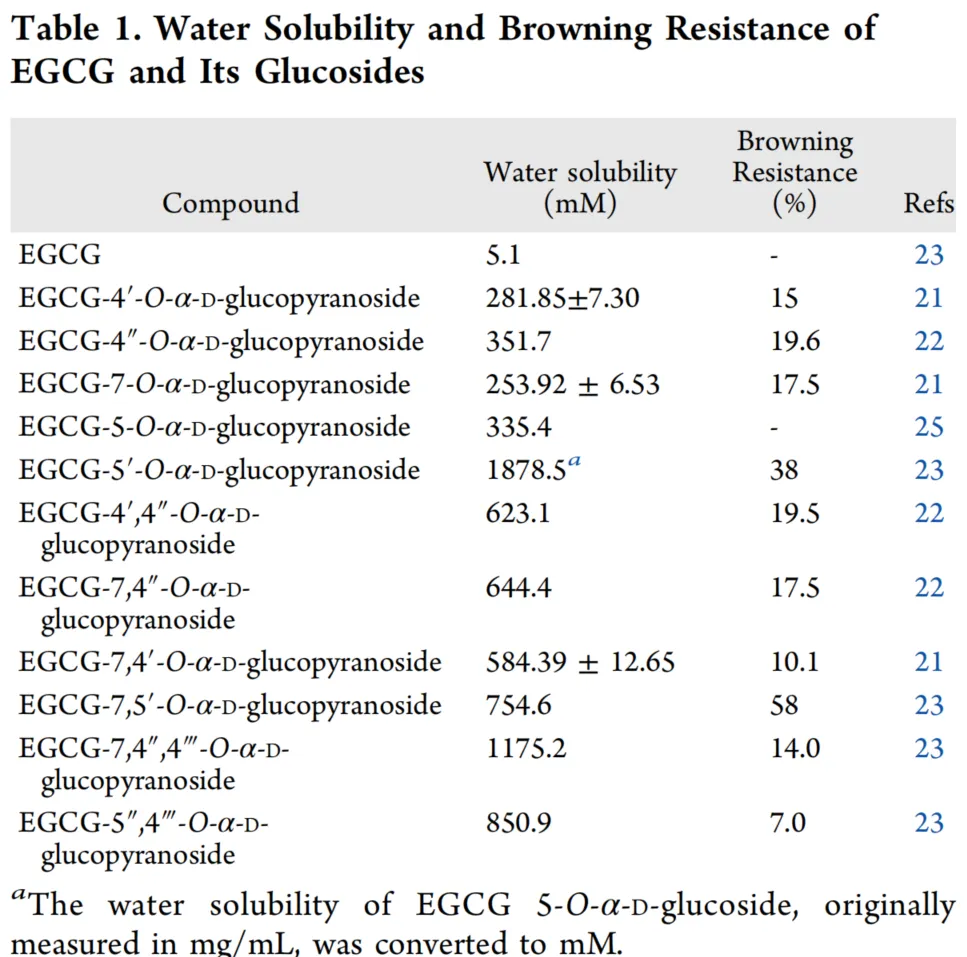
EGCG已通过甲基化、乙酰化、酯化、和糖基化进行修饰,以产生具有增强特性的衍生物。据报道,大多数具有生物活性的天然产物以糖苷形式存在,而糖基化具有增加化合物溶解度的显著优势。目前,糖基化主要通过化学方法和酶法实现。Zhang及其同事采用化学方法选择性生产了两种EGCG葡萄糖苷,证明这些糖苷比EGCG具有更高的稳定性和水溶性。尽管化学方法具有成本效益,但它们在后续纯化中带来挑战。相比之下,酶法具有高特异性、易于纯化和环境友好等优点。先前的研究报道了使用葡聚糖蔗糖酶、蔗糖磷酸化酶、环糊精糖基转移酶、和一种α-淀粉酶同源物(TRa2)生产EGCG α-葡萄糖苷。这些研究证实,EGCG葡萄糖苷比EGCG本身具有更高的水溶性和稳定性(表1)。然而,这些酶产生的EGCG葡萄糖苷相对多样,这使其应用复杂化。
蔗糖磷酸化酶是糖苷水解酶家族GH13的成员,催化蔗糖的磷酸解生成α-葡萄糖1-磷酸和果糖。它是一种多功能的转糖基酶,用于合成有价值的糖苷和糖。SPase存在于细菌中,如青春双歧杆菌(BiSPase)、肠膜明串珠菌(LmSPase)、嗜热杆菌(TSPase)和变形链球菌(SmSPase)。它已被用于催化天然产物的糖基化,包括黄酮类化合物。1995年,首次报道使用LmSPase生产了两种糖苷:(−)-表没食子儿茶素没食子酸酯4′-O-α-D-吡喃葡萄糖苷(EGCG-G1)和(−)-表没食子儿茶素没食子酸酯4′,4″-O-α-D-二吡喃葡萄糖苷(EGCG-2G)。最近,Chen等人使用SmSPase合成了EGCG-G1,并同时通过SPase的环工程阐明了EGCG糖基化所涉及的结构细节。显然,SPase是生产糖缀合物的一种非常有前景的生物催化剂;然而,其转糖基活性往往无法超越竞争性的水解反应。此外,工业生产中使用的SPase均来自非嗜热菌株,稳定性有限,这在一定程度上限制了其应用。因此,必须对SPase进行改造以增强其催化效率和耐热性。Gonzalez-Alfonso及其同事通过分子对接模拟分析生成了TtSPase的R134A突变体,分别实现了53%的根皮素单葡萄糖苷和73%的根皮素二葡萄糖苷的转化率。通过半理性诱变结合低通量筛选,开发了BiSPase双突变体(L341I_Q345S),实现了74%的曲二糖产率。通过应用半理性设计方法结合FireProt,研究人员创建了一个LmSPase突变体,其在50°C的半衰期几乎是野生型酶的两倍。
EGCG-G1是EGCG糖苷衍生物之一,与EGCG相比具有优越的性能,包括增强的水溶性、光稳定性和抗氧化活性。EGCG-G1最初是在LmSPase催化的EGCG糖基化产物中鉴定出来的。尽管LmSPase此前已被工程改造以提高其热稳定性,但其生产EGCG糖苷的能力尚未被重新评估。此外,较差的区域选择性仍然是阻碍LmSPase高效生产EGCG-G1的一个显著限制。在本研究中,工程化了LmSPase,以同时增强其活性、热稳定性、底物特异性和区域选择性,从而能够高纯度合成EGCG-G1,并尽量减少副产物。首先,在先前研究指导下进行的环工程产生了具有增强活性的L337A突变体。随后,共识设计产生了具有改善热稳定性的L337A/T219L双突变体。最后,通过两轮Rosetta Dock设计并结合有益突变,产生了具有显著增强的底物特异性和区域选择性的突变体M3(T219L/E393I/N335G)。这种整合策略成功地同时优化了多种酶学性质。在补料分批反应中使用突变体M3,实现了31.11 g/L EGCG-G1的生产,相对纯度为87.27%。
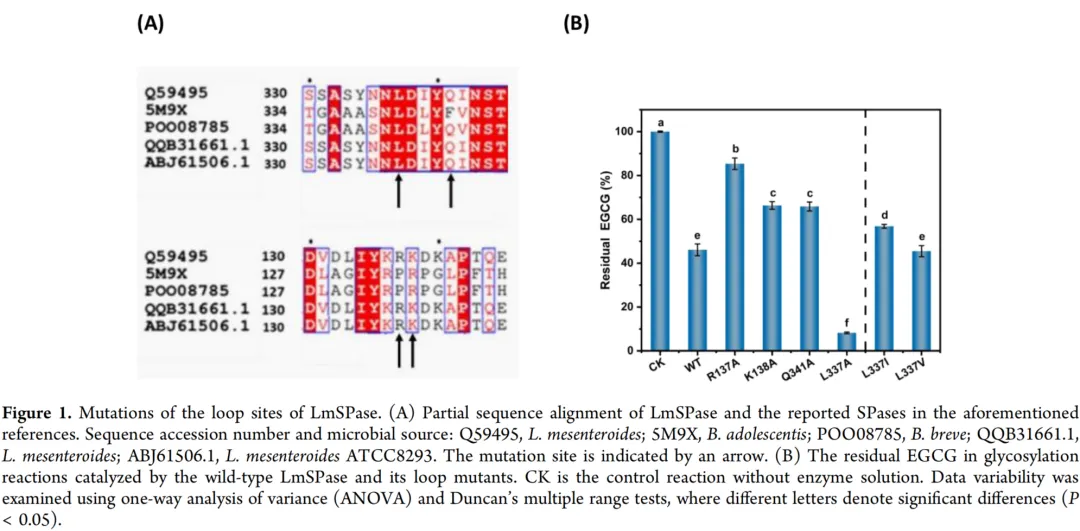
在BiSPase中,两个环——环A(残基336-344)和环B(残基130-140)——已被证明对底物特异性和催化效率有显著影响。当使用蔗糖磷酸化酶进行化合物糖基化时,野生型酶的催化效率往往达不到预期水平。由于这些环的柔性,对其氨基酸序列的修饰可以显著影响对底物的活性。因此,一些研究集中于SPase的环工程。例如,Ao及其同事对源自肠膜明串珠菌的SPase进行了饱和诱变,产生了一个显著的突变体R137F,其比活性比野生型酶高出约161%。环位点的突变可能会改变底物通道的大小,从而影响活性。来自肠膜明串珠菌ATCC 8293的SPase突变体K138C的活性比野生型酶高160%。结构分析表明,K138C是调节底物结合口袋的关键功能残基,从而影响活性。来自短双歧杆菌的SPase双突变体L341V/L343F几乎完全控制了对L-抗坏血酸2-OH基团的选择性生产,实现了超过99%的选择性。来自青春双歧杆菌的SPase的Q345F突变体对白藜芦醇、(+)-儿茶素和(−)-表儿茶素的糖基化效率高达97%。Febres-Molina等人研究了Q345F突变体催化的白藜芦醇糖基化的反应机理。
为了增强EGCG的糖基化能力,本研究首先对位于LmSPase环区的四个特定位点(R137、K138、L337和Q341)进行了丙氨酸扫描。在这些位点中,L337和Q341分别对应于短双歧杆菌(POO08785)和青春双歧杆菌(5M9X)SPase中的L341和Q345(图1A)。使用野生型LmSPase及其突变体在pH 6.0和30°C下对EGCG(10 g/L)进行糖基化反应20小时。然后使用HPLC测量剩余的底物水平。结果表明,突变体R137A、K138A和Q341A反应混合物中的残留EGCG高于野生型,而突变体L337A的残留量小于10%(图1B)。为了研究其他潜在的氨基酸替换,考虑了先前的发现,即用异亮氨酸或缬氨酸替换BbSPase(POO08785)第341位的亮氨酸可使其对L-抗坏血酸的活性提高20%至47%。在LmSPase的L337位点引入了相同的突变,并评估了它们相对于野生型酶和L337A突变体的性能。与在BbSPase中观察到的结果不一致,LmSPase的L337I和L337V突变体没有表现出对EGCG的活性改善(图1B)。然而,L337A突变体实现了约90%的EGCG转化。随后对该单点突变体和野生型之间的转糖基酶活性(使用纯化酶)的比较显示,L337A突变体的转糖基酶活性(12.20 mU/mg)约为野生型(3.49 mU/mg)的3.50倍。
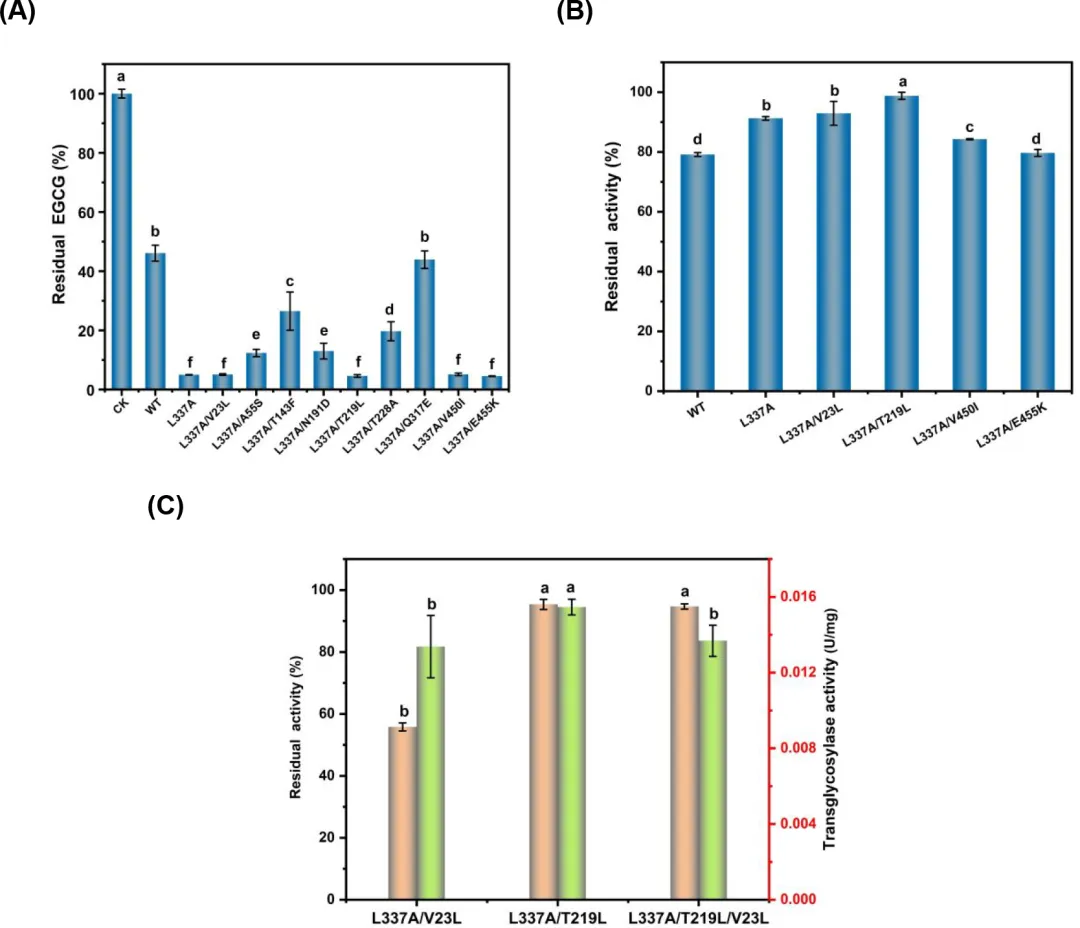
共识设计为创建既高度稳定又具有生物活性的蛋白质提供了一种有价值的方法。在评估野生型和突变体L337A的热稳定性时,它们在45°C下的半衰期分别为43.32分钟和49.51分钟。显然,L337A突变体对于长时间反应来说稳定性不足。将突变体L337A的序列提交给Consensus Finder以预测潜在的稳定化突变。结果识别出九个突变:T219L、V450I、V23L、Q317E、N191D、T228A、E455K、A55S和T143F。随后,创建了这些单点突变体并在大肠杆菌BL21(DE3)中表达。
在使用上述突变体进行EGCG糖基化时,在30°C反应20小时后,在四个突变体L337A/V23L、L337A/T219L、L337A/V450I和L337A/E455K的反应混合物中检测到的残留EGCG与L337A观察到的水平一样低(图2A)。在40°C孵育1小时后,突变体L337A/V23L和L337A/T219L的粗酶提取物中的剩余活性均超过野生型和L337A突变体,其中L337A/T219L表现出最高的剩余活性(图2B)。然后通过组合这三个有利突变创建了三重突变体L337A/V23L/T219L。该突变体表现出与L337A/T219L相当的热稳定性,在45°C孵育1小时后保持超过90%的活性。相比之下,突变体L337A/V23L在相同条件下保持不到60%的活性(图2C)。当使用纯化酶测量转糖基酶活性时,L337A/T219L突变体在三个比较的突变体中实现了最高的活性(图2C)。此外,测定L337A/T219L突变体的半衰期为6.54小时,分别是野生型和L337A突变体的9.05倍和7.93倍。如先前报道,通过半理性设计结合FireProt获得的T219L和I31F/T219L/T263L/S360A(MUT4)突变体在50°C下的半衰期约为野生型LmSPase的两倍。38 因此,很明显T219L突变对增强LmSPase的稳定性有积极作用。
3. 通过Rosetta Dock设计提高底物特异性和区域选择性如先前报道,LmSPase催化从蔗糖到EGCG的转葡萄糖基反应,产生两种稳定的EGCG葡萄糖苷:EGCG-G1和EGCG-2G。在本研究中,EGCG-G1被鉴定为野生型LmSPase催化的EGCG糖基化的主要产物。尽管双位点突变体L337A/T219L表现出增强的活性和稳定性,但它仍然产生EGCG葡萄糖苷的混合物。除了EGCG-G1和EGCG-2G外,通过HPLC-MS还鉴定出另一种次要的单葡萄糖苷,显示[M−H]⁻离子在m/z 619.1305处。该化合物推测为(−)-表没食子儿茶素没食子酸酯4″-O-α-D-吡喃葡萄糖苷,并在本研究中命名为EGCG-G2。产物的复杂性使得EGCG-G1的分离复杂化,从而限制了其应用。
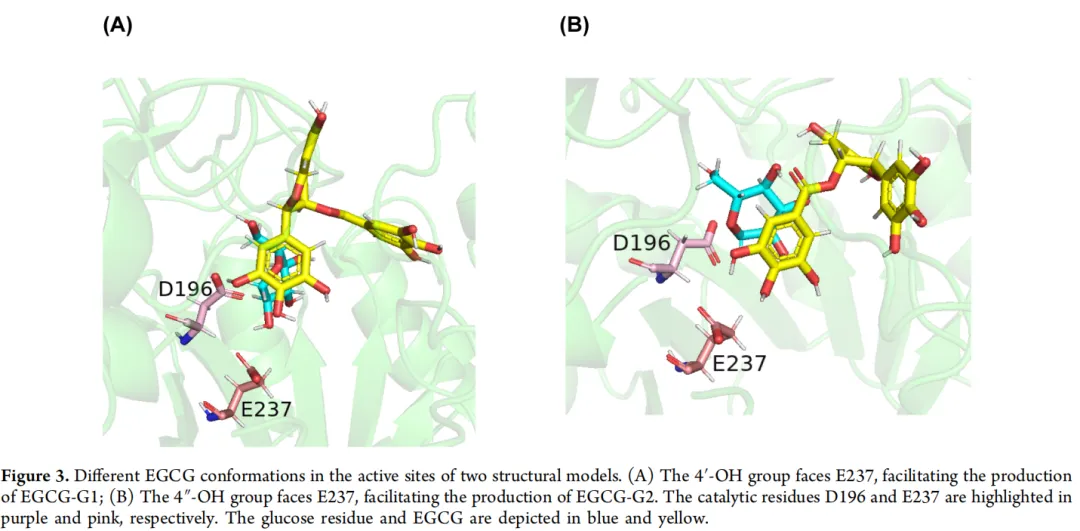
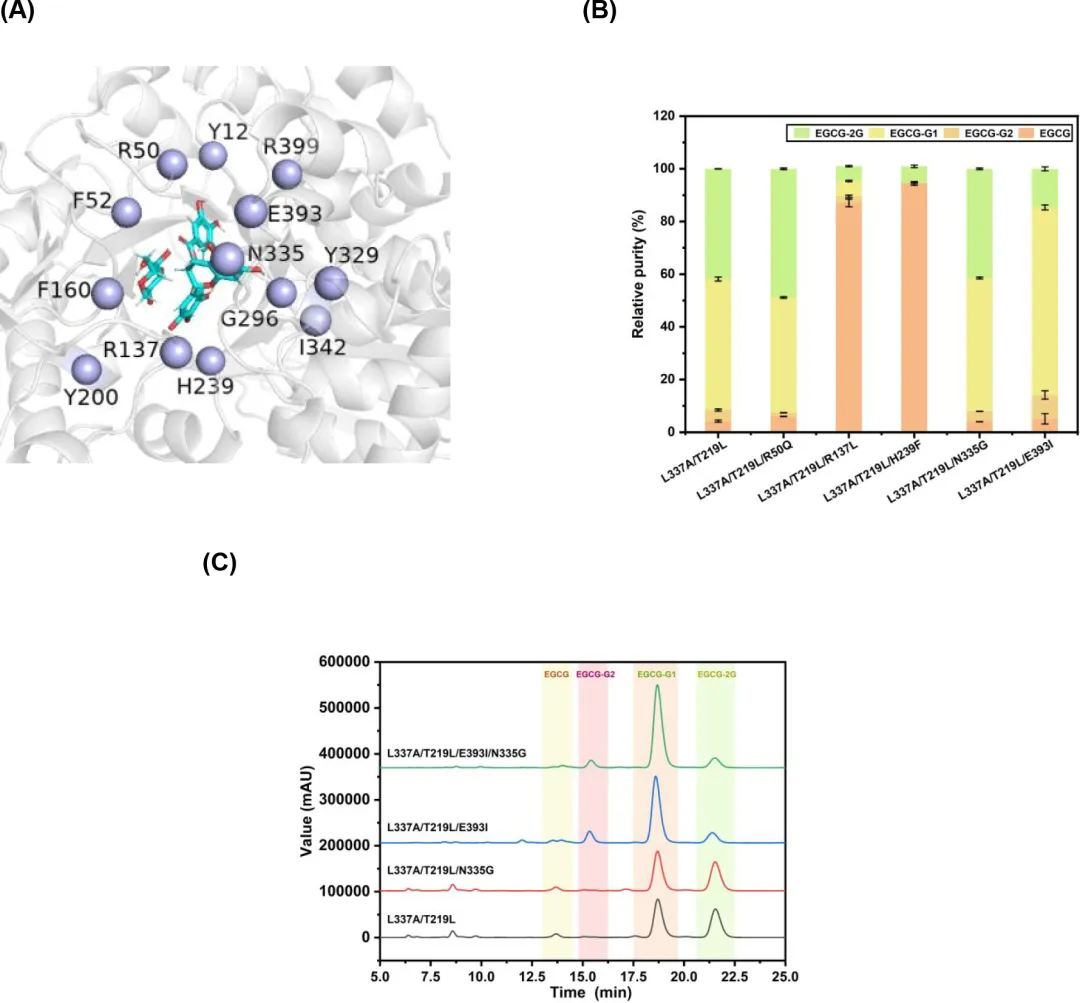
为了获得具有改进底物特异性和区域选择性的LmSPase突变体,使用Rosetta Dock设计来修饰活性位点内的蛋白质-配体界面。在准备阶段,选择了与葡萄糖基和EGCG复合的L337A/T219L突变体的两个结构模型。这些模型表现出不同的构象,其中4'-OH和4″-OH基团朝向E237,分别促进EGCG-G1和EGCG-G2的产生(图3)。随后使用MD模拟对这些复合物结构进行了细化。总共选择了位于4'-OH或4″-OH基团附近且在EGCG 5 Å范围内的13个残基(Y12、R50、F52、F160、R137、Y200、H239、G296、Y329、N335、I342、E393和R399)(图4A),并按照Rosetta对接和设计方案将其突变为所有19种氨基酸。从生成的5000个模型中,根据它们的总能量、界面能量得分以及配体和蛋白质之间的距离选择了25个结构。然后收集并对齐相应的序列,揭示了十个具有高频突变的位点(Y12、R50、F160、R137、Y200、H239、Y329、N335、E393和R399)。对于每个位点,使用FoldX分析了比对结果中出现的突变氨基酸。将五个ΔΔG < 0的突变(R50Q、R137L、H239F、N335G和E393I)分别引入L337A/T219L突变体,并在20小时的EGCG糖基化反应中进行实验验证(图4B)。与双位点突变体L337A/T219L相比,由三位点突变体L337A/T219L/E393I催化的反应在EGCG葡萄糖苷中显示出更高比例的EGCG-G1,产生了7.55 g/L(12.17 mmol/L)的EGCG-G1,表明对EGCG-G1的区域选择性有所提高。引入R50Q突变(L337A/T219L/R50Q)导致EGCG-2G的产量增加,而包括R137L和H239F在内的其他突变(L337A/T219L/R137L和L337A/T219L/H239F)导致活性完全丧失。N335G突变似乎没有损害酶活性或特异性,因此将其引入L337A/T219L/E393I突变体,产生了四位点突变体L337A/T219L/E393I/N335G(M4)。经证实,在含有突变体M4的20小时反应混合物中,与L337A/T219L/E393I相比,EGCG-G1的相对纯度有所提高(图4C),约占总产物的81.98%,产生了9.47 g/L(15.27 mmol/L)的EGCG-G1。
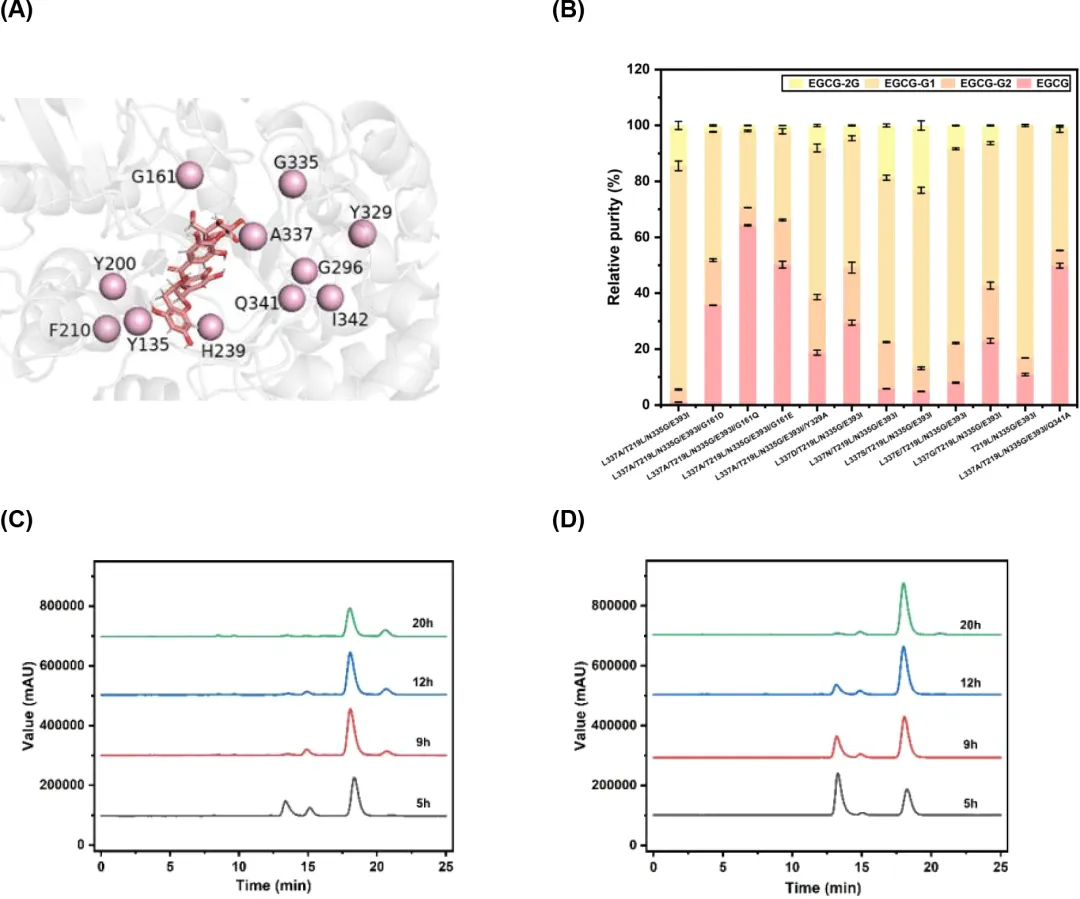
为了进一步提高突变体M4的特异性和区域选择性,使用RosettaLigand生成了其复合物模型,随后通过MD模拟进行细化。在选择突变位点时,我们避免了有利于4″-葡萄糖苷形成的突变。因此,选择残基Y135、G161、Y200、F210、H239、G296、Y329、G335、A337、Q341和I342进行重新设计(图5A)。同样,选择了83个突变体结构,并收集其序列进行比对。最后,选择了25个突变体进行实验验证,包括Y135(A/D/G/N/S/V)、F210H、Y200G、H239(F/G/V/W)、Q341(L/T/A)、G161(D/E/Q)、Y329A和A337(L/D/E/G/N/S)。EGCG糖基化反应的结果表明,当将前述突变位点分别引入M4时,突变Y135(A/D/G/N/S/V)、F210H、Y200G、H239(F/G/V/W)和Q341(L/T)完全消除了转糖基酶活性。相比之下,突变G161(D/E/Q)、Y329A、A337(D/E/G/N/S/)和Q341A相对于M4导致区域选择性降低。M4的A337L突变体在产生EGCG-G1方面获得了增强的特异性和区域选择性。在30°C反应20小时后,产生的EGCG-2G极少,且EGCG-G1的产量超过了原始的M4突变体。特别地,当第337位残基突变为亮氨酸时,突变体T219L/E393I/N335G(M3)表现出约占总产物83.18%的EGCG-G1比例,产生了9.56 g/L(15.42 mmol/L)的EGCG-G1,产率为70.61%(图5B),表明区域选择性显著提高。HPLC分析显示,突变体M4催化的反应比突变体M3催化的反应显示出更快的糖基化速率。M4在大约9小时内转化EGCG,而M3的转化需要超过12小时(图5C和D)。此外,在M4催化的反应5小时后开始形成EGCG-2G,并且其数量随着反应的进行稳步增加(图5C)。相比之下,即使在20小时后,M3催化的反应也只产生少量的EGCG-2G,表明具有更高的特异性和区域选择性(图5D)。
最后,评估了L337A/T219L(6.54小时)、L337A/T219L/E393I(2.77小时)、M4(2.68小时)和M3(1.94小时)的半衰期。尽管与L337A/T219L相比,后三个突变体的稳定性有所下降,但考虑到在24小时反应中EGCG-G1的积累,它们增强的活性和区域选择性足以弥补这一损失。因此,表现出最佳区域选择性的M3被确定为最佳突变体。突变体M3展现出显著提高的催化效率和热稳定性。其在30°C下的转糖基酶活性达到17.77 mU/mg,是野生型(3.49 mU/mg)的4.09倍。同时,其在45°C下的半衰期相对于野生型延长了1.69倍。纯化了野生型和突变体M3,并测定了它们对EGCG的动力学参数,包括Km、kcat和kcat/Km。对于野生型,这些值分别为0.29 ± 0.02 mM、0.02 ± 0.00 s⁻¹和0.07 mM⁻¹ s⁻¹;对于突变体M3,它们分别为0.28 ± 0.03 mM、0.04 ± 0.00 s⁻¹和0.14 mM⁻¹ s⁻¹。正如Km值所示,野生型和M3对EGCG表现出相似的结合亲和力;然而,M3对EGCG表现出更高的kcat和更高的kcat/Km值(约2倍)。


结构分析表明,位于活性位点内的335、337和393位点的突变可能会改变LmSPase底物结合口袋的形状。分别用G335、A337和I393替换N335、L337和E393引入了较小的氨基酸侧链,从而减少了底物进入催化口袋的空间位阻并影响底物结合。这种修饰增强了活性并在催化过程中调节底物特异性。类似地,Duan等人将肠膜明串珠菌ATCC 8293的SPase第138位的赖氨酸残基替换为侧链较短的半胱氨酸,从而暴露了催化口袋。这种修饰促进了底物进入活性位点,从而增加了活性。在早期的实验验证中,观察到将第337位的亮氨酸突变为丙氨酸可增强活性(图1B)。由于第337位残基位于LmSPase的环A中,该环显著影响活性,推测第337位点主要有助于活性的增加,而第335和393位点主要影响特异性的改善。然而,在第二轮设计中,观察到与突变体M4(L337A/T219L/E393I/N335G)相比,在突变体M3(T219L/E393I/N335G)中将A337恢复为L337并没有降低活性或特异性。事实上,其特异性相对于M4有所增加,并且几乎不产生EGCG-2G。因此,第219、335和393位的残基可能协同作用以增强活性和特异性,使突变体M3能够高效生产EGCG-G1。第219位残基位于远离活性位点的位置。CavityPlus分析表明,N335G和E393I突变导致底物结合口袋的形状和体积发生变化,从而使底物与突变体M3的活性位点之间更加契合。
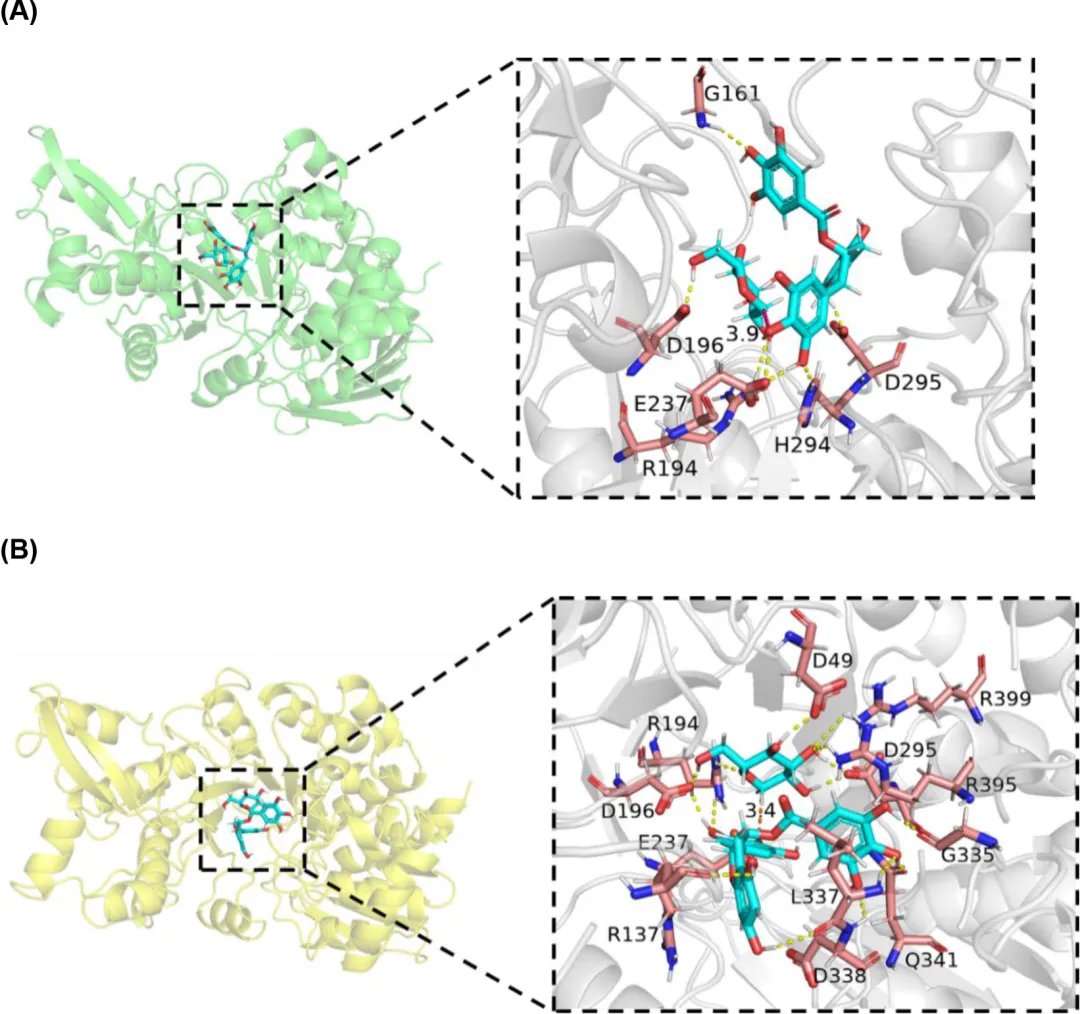
在50 ns MD模拟中复合物的RMSD显示,M3的结构比野生型更稳定。疏水相互作用和氢键是蛋白质热稳定性的关键贡献因素。通过整合在催化口袋内部或附近形成疏水相互作用和氢键的残基,理解它们在活性位点配体结合中的额外作用对于同时增强热稳定性和活性至关重要。在野生型LmSPase中,葡萄糖基与R194和D196形成氢键,而EGCG通过与G161、E237、H294和D295的氢键相互作用(图6A)。在突变体M3中,葡萄糖基与D49、R194、D196、D295、R395和R399建立氢键,而EGCG与R137、E237、G335、L337、D338和Q341形成氢键(图6B)。这些观察结果表明,在M3突变体中,葡萄糖基和EGCG与活性位点中氨基酸残基之间的氢键相互作用都增强了。同时,葡萄糖基与D295形成氢键,有助于稳定LmSPase催化的葡萄糖基转移反应过程中的中间体。对于EGCG,G335和L337与含有4'-OH的苯环上的羟基形成氢键。具体来说,G335、L337和D338分别与3″-OH、4″-OH和5″-OH结合。因此,它们阻断了葡萄糖基的结合位点,从而增强了特异性和区域选择性。这可以解释为什么当A337突变恢复为L337时反应特异性增加。此外,对整个MD模拟过程中葡萄糖基与4'-OH和4″-OH基团之间距离变化的分析显示,葡萄糖基通常保持更靠近4'-OH基团。比较野生型和M3突变体的代表性结构显示,葡萄糖基与EGCG的4'-OH基团之间的距离从3.9 Å减小到3.4 Å(图6)。糖基供体和受体之间更短的距离可能会增加活性。62 此外,口袋疏水性的增加可能增强其对疏水性底物的可及性,从而加强活性位点与底物之间的结合。因此,这些氢键和疏水相互作用很可能都有助于M3突变体稳定性、活性和特异性的提高。
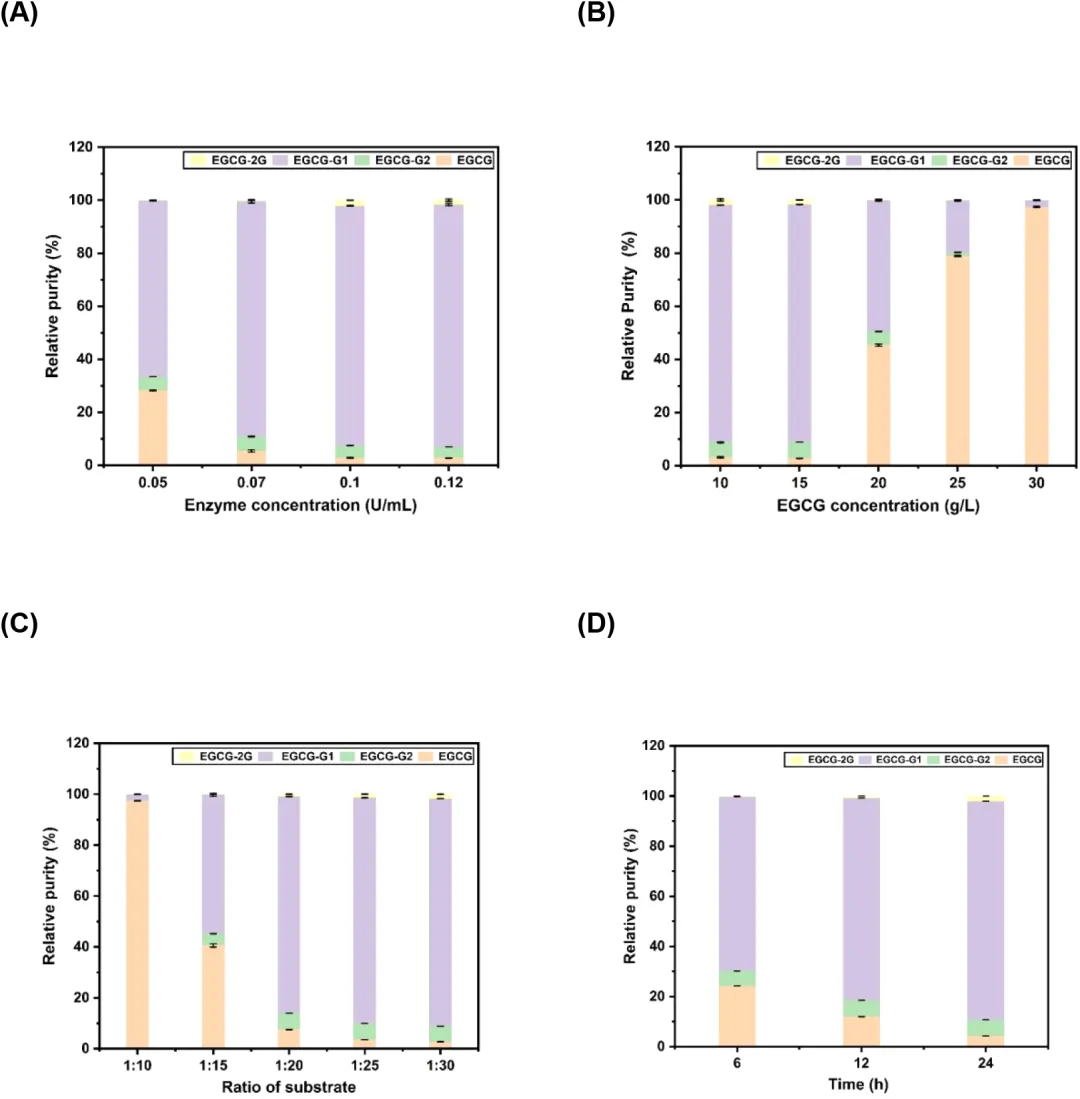
5. 利用LmSPase突变体高效生产EGCG-G1使用T219L/N335G/E393I突变体(M3)的粗酶,优化了反应条件以高效生产EGCG-G1。研究了影响因素,包括反应时间、酶浓度、底物浓度和底物比例。测试了四种酶浓度(0.05、0.07、0.1和0.12 U/mL),在反应过程中于3、5、7、9和12小时取样。观察到随着酶浓度逐渐增加,EGCG的消耗速率相应加快。在酶浓度为0.05或0.07 U/mL时,EGCG在12小时内完全消耗,HPLC无法检测到。在酶浓度为0.1或0.12 U/mL时,EGCG在7小时内完全转化(图7A)。因此,在0.12 U/mL的酶浓度下,增加EGCG浓度进行9小时的反应。如图7B所示,15 g/L的EGCG也完全转化。接下来,当初始EGCG浓度为15 g/L时,调整EGCG与蔗糖的摩尔比。在1:30的比例下,EGCG-G1的相对纯度达到最高水平(图7C)。在最佳条件下,包含15 g/L EGCG、336 g/L蔗糖(摩尔比1:30)和0.12 U/mL粗酶,在30°C下反应9小时,产生15.23 g/L(24.57 mmol/L,产率75.01%)的EGCG-G1,通过HPLC测定相对纯度为89.47%。为了转化更多的EGCG,采用了补料分批策略。在5 mL反应混合物中,初始底物浓度为15 g/L,EGCG与蔗糖的摩尔比为1:30,温度为30°C,反应6小时后加入5 g/L EGCG。再过6小时后,再加入5 g/L EGCG,并在30°C下继续反应12小时。如图7D所示,15 g/L的EGCG在6小时内未完全转化;然而,当补充额外的5 g/L EGCG并在12小时取样时,剩余的EGCG浓度甚至低于6小时时的浓度,表明这种LmSPase突变体具有鲁棒性。最终,在24小时内总共转化了25 g/L EGCG,产生了31.11 g/L(50.18 mmol/L)的EGCG-G1,相对纯度为87.27%,总产率为91.92%。这项工作开发了一种能够高效且特异性生产EGCG-G1的LmSPase突变体,并为EGCG葡萄糖苷的大规模生产奠定了基础。