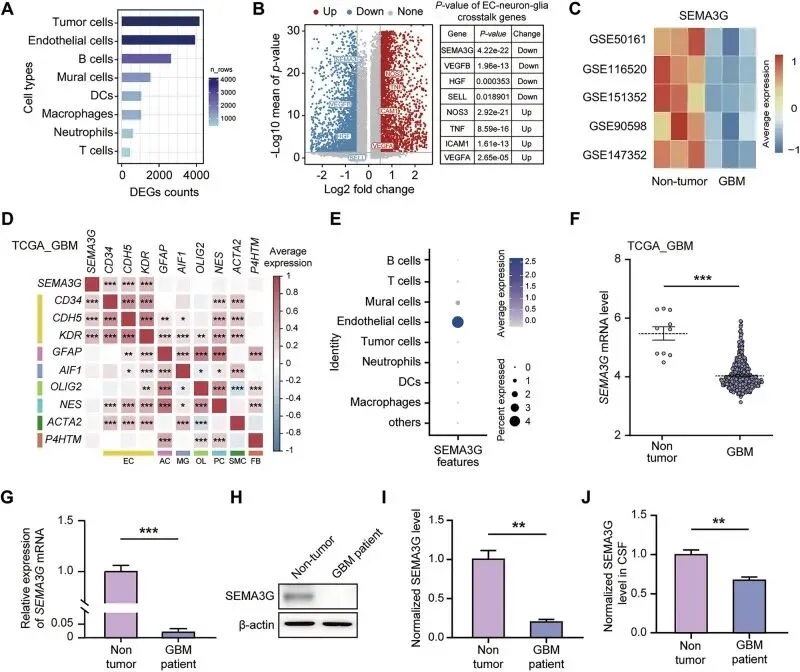
SEMA3G在GBM患者中低表达,且主要源于内皮细胞
直接锁定了SEMA3G这个靶点。从单细胞数据(GSE162631)一看,GBM内皮细胞里它掉得最狠,P值极显著。后续在多中心数据库和临床样本里都得到验证:不管是mRNA还是蛋白,肿瘤里都比正常脑组织和脑脊液低一大截,而且它和内皮标志物CD34/CDH5完全同步,实锤主要由内皮细胞分泌。
复现建议:
如果你的课题也关注某种分泌蛋白在肿瘤微环境中的作用,可以借鉴这套“单细胞数据初筛 -> 多数据库验证 -> 自有样本qPCR/WB验证 -> 体液ELISA验证”的完整流程。在自己论文中,可以先用公共数据库快速锁定目标,再聚焦验证。
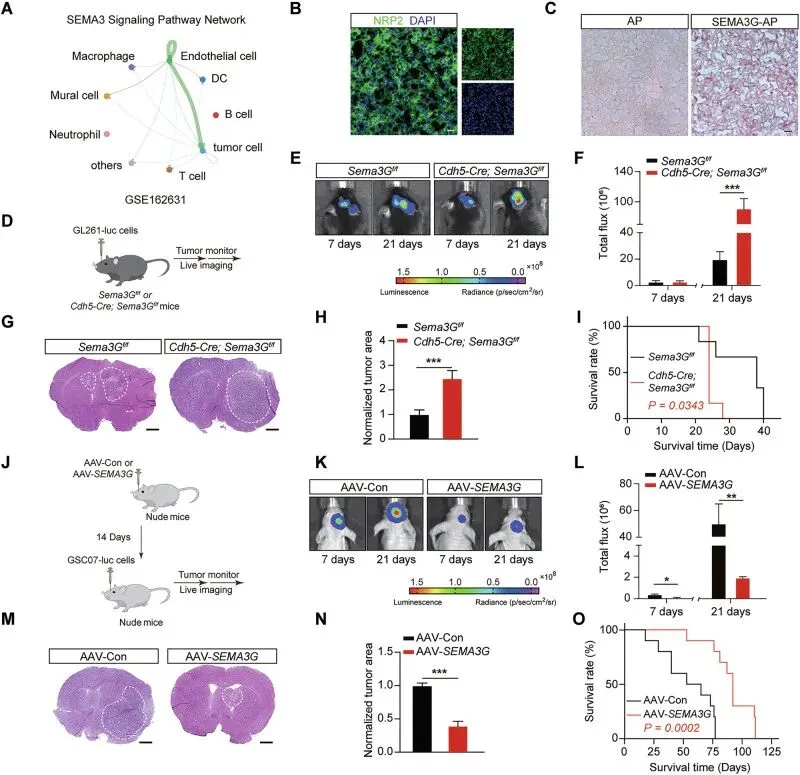
通过CellChat分析预测,SEMA3G的主要作用对象是肿瘤细胞(图2A),并通过免疫荧光和AP配体结合实验,证实SEMA3G确实能结合到GBM组织上,且依赖于其受体NRP2(图2B, C)。
为了验证功能,他们构建了内皮细胞特异性敲除Sema3G的小鼠。原位接种肿瘤后,敲除鼠脑内的肿瘤长得更快、更大,老鼠死得更早(图2D-I)。
复现建议:
这里非常推荐大家学他们这套“细胞特异性基因操作”的思路,做机制验证时特别管用。首选肯定是Cre-LoxP条件性敲除小鼠,直接在源头把目标细胞的功能“减”掉,干净利落;如果造小鼠周期太长,也可以用AAV局部注射或者腹腔打重组蛋白来做“加法”回补。
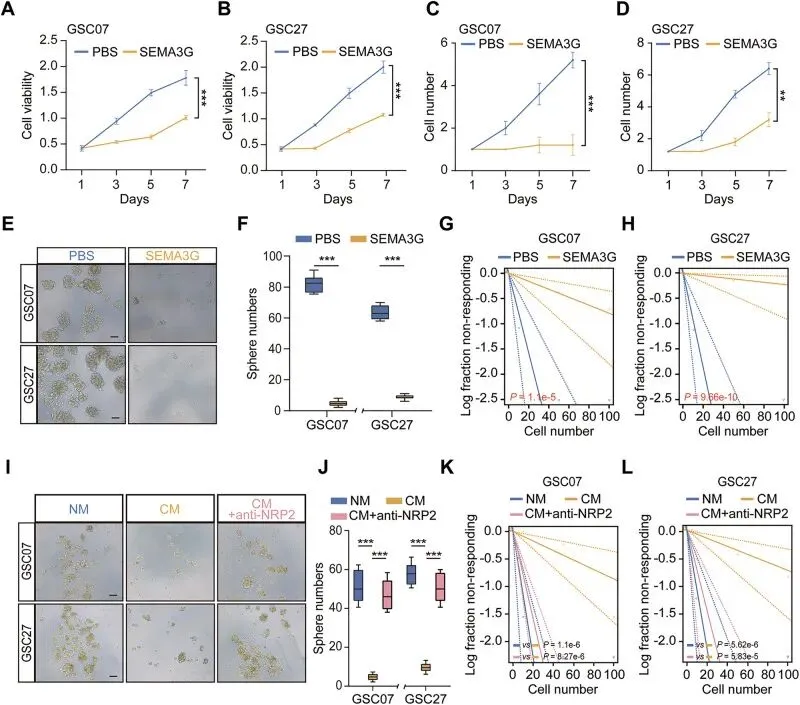
体外实验表明,重组SEMA3G蛋白能时间和浓度依赖性地抑制GSC的生长、神经球形成能力和自我更新能力(图3A-H)。更重要的是,他们收集了过表达SEMA3G的HUVEC细胞的条件培养基(CM) 来模拟内皮细胞的旁分泌作用。结果发现,这种CM同样能抑制GSC的干性,而这种抑制作用可以被NRP2的中和抗体所“挽救”(图3I-L)。
复现建议:
强烈推荐使用Transwell共培养或条件培养基体系。设计实验时,务必设置“挽救”组,这能让你的机制研究从“相关性”走向“因果性”。
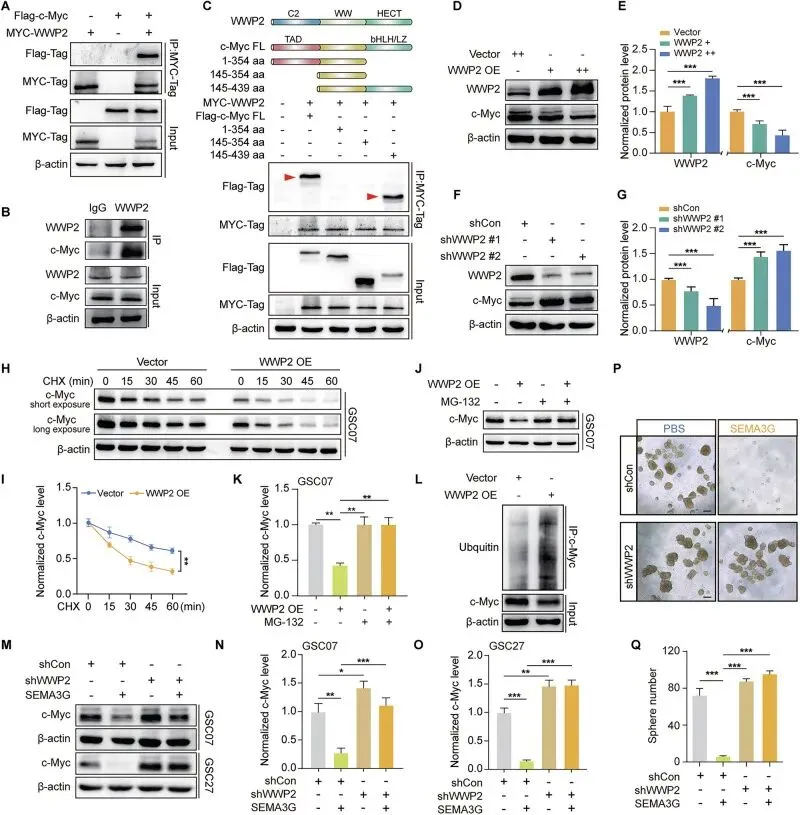
SEMA3G通过泛素-蛋白酶体途径促进c-Myc降解
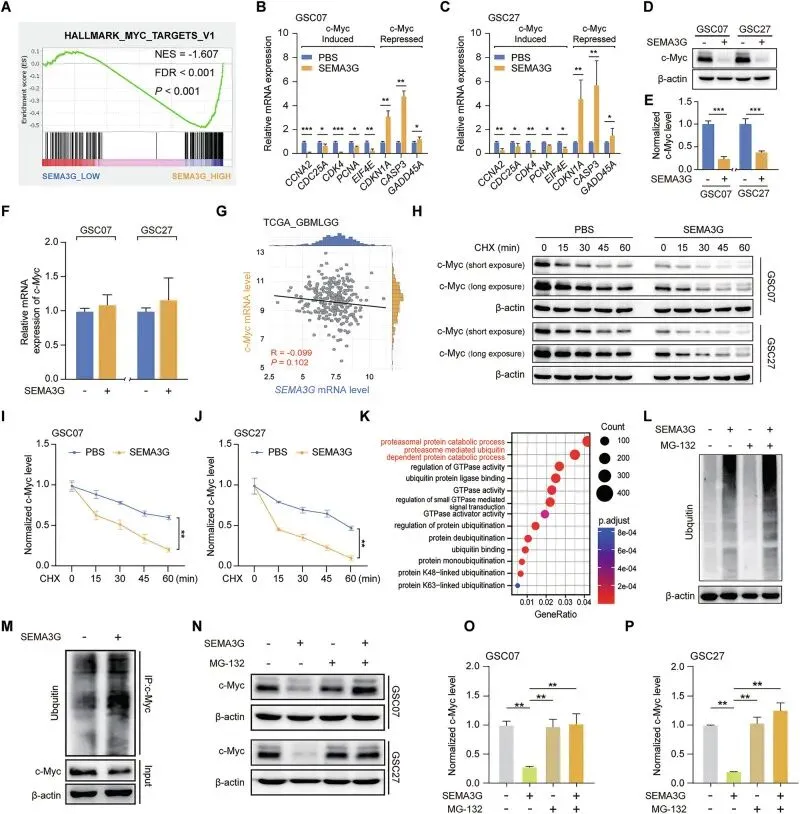
把SEMA3G怎么干掉c-Myc的机制摸透了。GSEA一看,SEMA3G低的病人,c-Myc通路疯涨。说明是翻译后调控。放线菌酮实验显示c-Myc的半衰期明显变短了。实验证实SEMA3G只管蛋白不管RNA,蛋白半衰期实验更是实锤:加了SEMA3G,c-Myc死得快。最后锁定是泛素化途径,SEMA3G能拉泛素化给c-Myc贴“死亡标签”,用MG-132一堵,蛋白就不降了,逻辑链完美闭环。
图4.SEMA3G在GSCs中降低c-Myc蛋白的稳定性
复现建议:
当发现一个蛋白水平变化但mRNA不变时,CHX追踪半衰期和MG-132处理是必须做的两个实验。在自己的论文里,如果发现类似现象,可以立刻沿着“泛素化”这条路往下走,去筛选E3连接酶或去泛素化酶。
WWP2是SEMA3G诱导c-Myc降解的关键E3泛素连接酶
抓到了真正的“刽子手”WWP2。作者先筛了6个候选E3连接酶,Co-IP一筛,只有WWP2能跟c-Myc抱团。截短体实验精准定位到了c-Myc的bHLH/LZ区域是结合位点。功能上,WWP2就是降解开关:过表达它就降,敲低它就升。最绝的是在GSC里敲掉WWP2,SEMA3G彻底失效,既降不了c-Myc,也干不掉神经球,实锤SEMA3G全靠WWP2干活。
图5.SEMA3G通过依赖WWP2的方式促进c-Myc降解
复现建议:
这里强烈推荐大家抄这套“互作筛选+功能挽救”的作业,做机制的时候特别顺手。找互作蛋白别瞎蒙,先跑个质谱或者翻翻BioGRID、STRING这些数据库,圈几个候选出来再用Co-IP去验证,命中率能高不少。
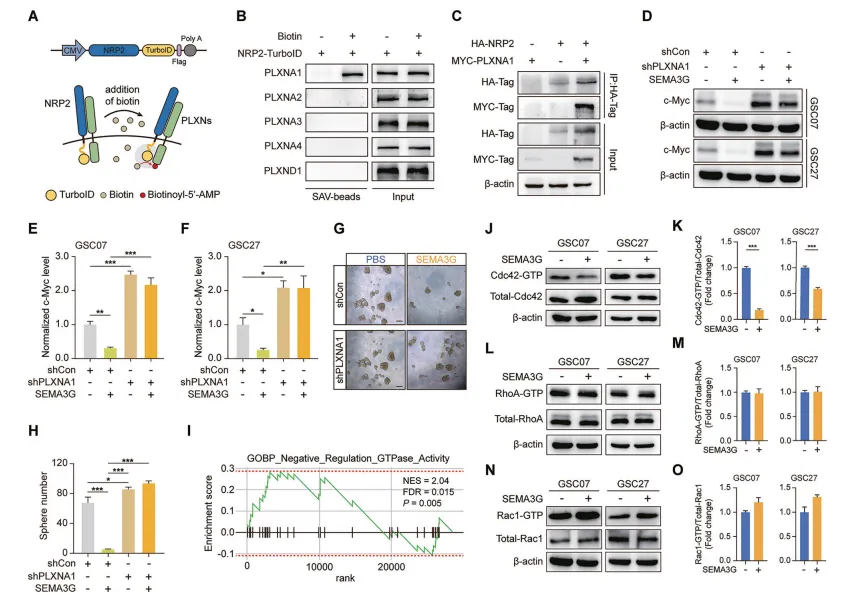
SEMA3G通过NRP2/PLXNA1受体复合物失活Cdc42
通过TurboID邻近标记和Co-IP,确定SEMA3G的受体复合物是NRP2/PLXNA1(图6A-C)。敲低PLXNA1同样能阻断SEMA3G对c-Myc的下调作用(图6D-H)。
GO分析提示“GTPase活性负调控”。通过GST-Pull-down实验,他们发现SEMA3G特异性地抑制了Cdc42的活性,而对RhoA和Rac1影响不大(图6I-O)。
图6.SEMA3G通过GSCs中的NRP2/PLXNA1失活Cdc42
复现建议:
推荐大家学这套“互作+活性”的组合拳,做信号通路机制时特别管用。想找受体复合物,普通实验室没必要死磕高门槛的TurboID,踏踏实实用Co-IP加免疫荧光共定位就是最稳的黄金搭档。
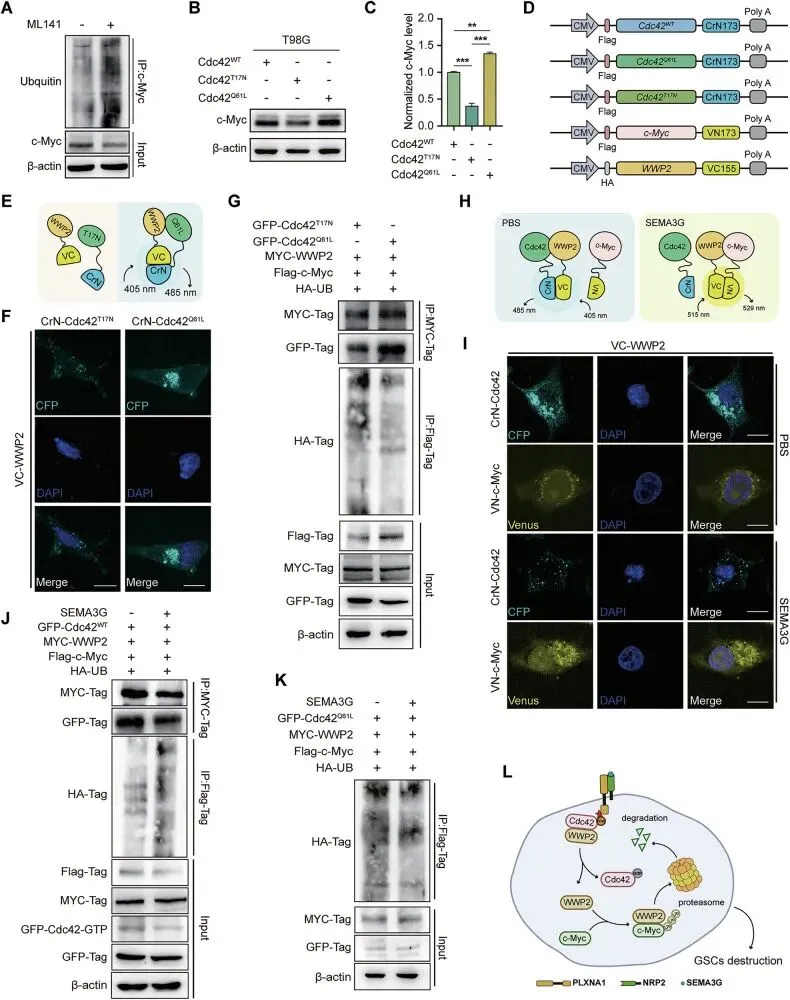
Cdc42充当分子开关,调控WWP2与c-Myc的结合
把这台“分子开关”拆得明明白白。用Cdc42的突变体一看:死掉的Cdc42(T17N)能让c-Myc跟着陪葬,而活蹦乱跳的Cdc42(Q61L)则把它护得死死的。机制上更精妙,平时Cdc42像个“保镖”一样把WWP2扣住,不让它碰c-Myc;一旦SEMA3G来了,Cdc42失活(GDP态),WWP2立马被释放,转身就把c-Myc拖去降解了。
图7.Cdc42充当分子开关,调控WWP2介导的c-Myc降解过程
复现建议:
研究这种复杂的动态竞争机制,BiFC或FRET技术是很好的选择。在研究中,一个蛋白的非经典功能可能比其经典功能(GTPase)更重要。在自己的课题中,当发现两个蛋白功能相关但无直接催化关系时,可以大胆假设存在一个“竞争性结合”或“扣押-释放”的模型。