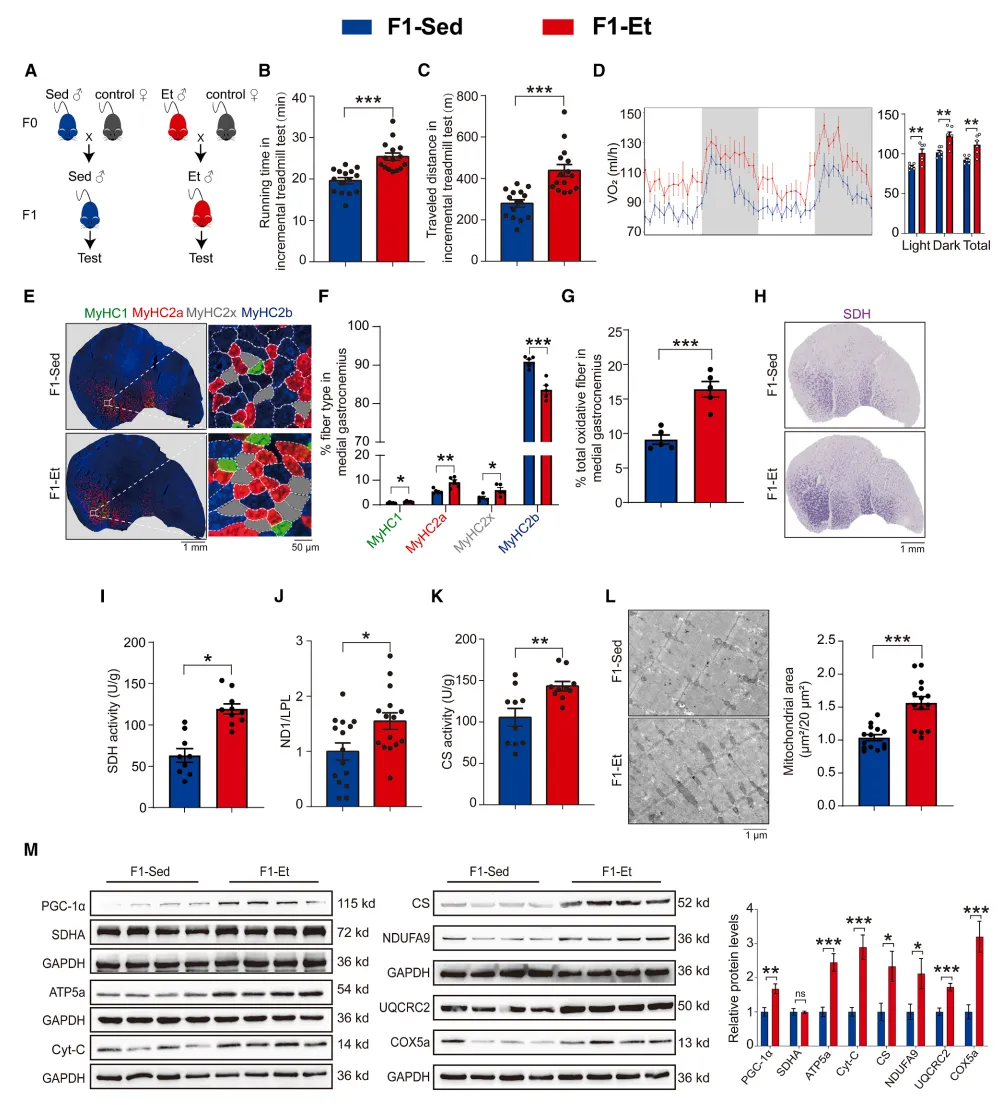
确立父代运动可跨代增强子代耐力与骨骼肌氧化代谢的表型
研究首先给出了最直观的结论:坚持运动的雄性小鼠,生下的后代即便从小不运动,天生耐力、有氧代谢能力也显著优于久坐父本的子代。背后是骨骼肌从细胞层面发生了永久性重塑:肌纤维更多转向耐疲劳、高氧化型,线粒体数量、合成能力、功能活性全面升级。相当于父亲提前通过表观遗传,给孩子的骨骼肌 “预装” 了高配运动体质【图1】。
图1.父代运动可提升子一代的耐力水平,促使肌纤维类型发生转变,并增加其线粒体数量
小编总结:父代运动不是只利己,而是直接给后代骨骼肌埋下高耐力、高代谢的先天生理基础。
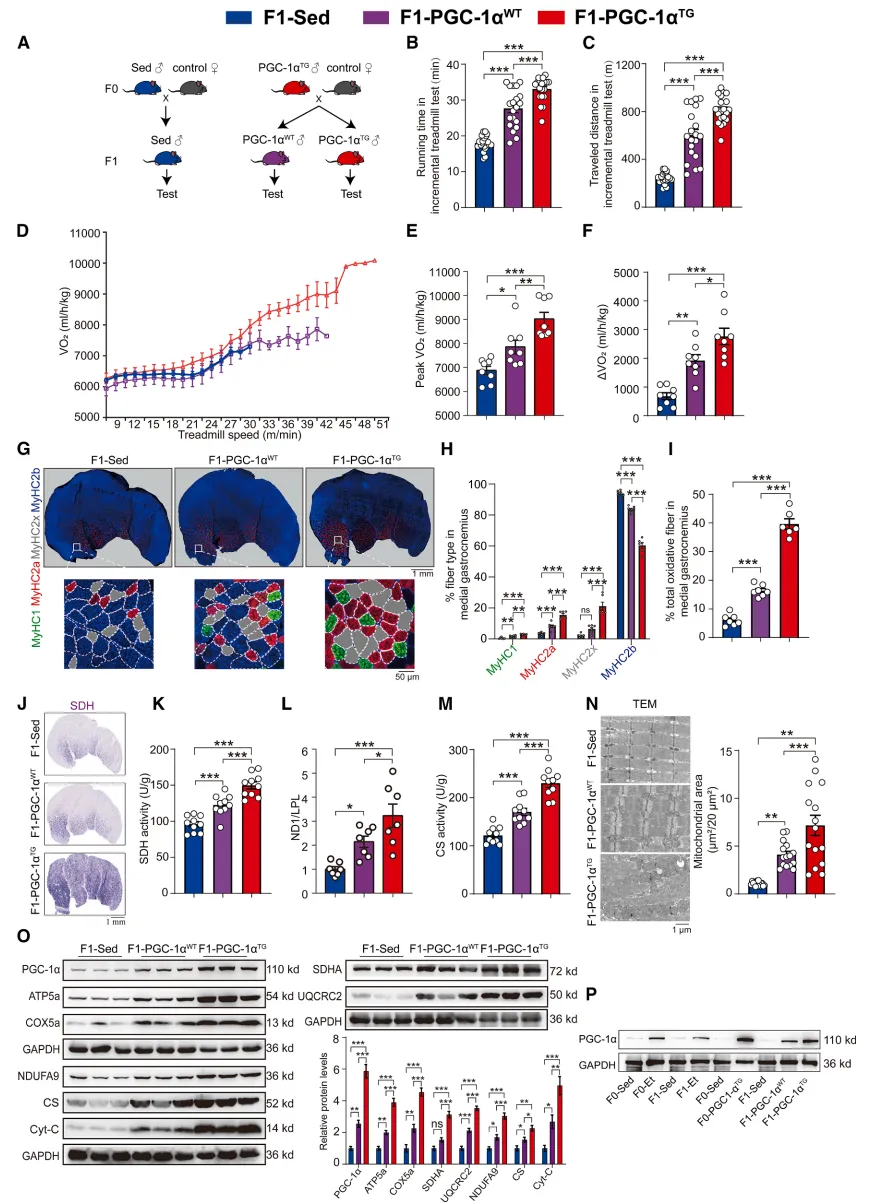
利用转基因模型证明父代肌肉PGC-1α激活足以触发跨代遗传
为了证明不是运动行为本身,而是肌肉代谢状态决定跨代遗传,研究用到了转基因模型:只在父亲骨骼肌里强行激活线粒体核心基因PGC-1α,不需要任何跑步训练,依然能让不携带转基因的子代,复刻出高耐力、肌纤维转型、线粒体增多的同款表型。这一步直接把上游关键因子锁定:父代肌肉PGC-1α激活,是触发运动获益跨代传递的核心开关【图2】。
图2.在首代转基因小鼠中实现肌细胞特异性过表达PGC-1α,可提升其耐力,并促使未携带转基因的子一代小鼠发生肌纤维类型转换与线粒体生物合成
小编总结:真正负责传代的是肌肉代谢核心通路激活,而非跑步动作本身,精准找到了上游调控关键。
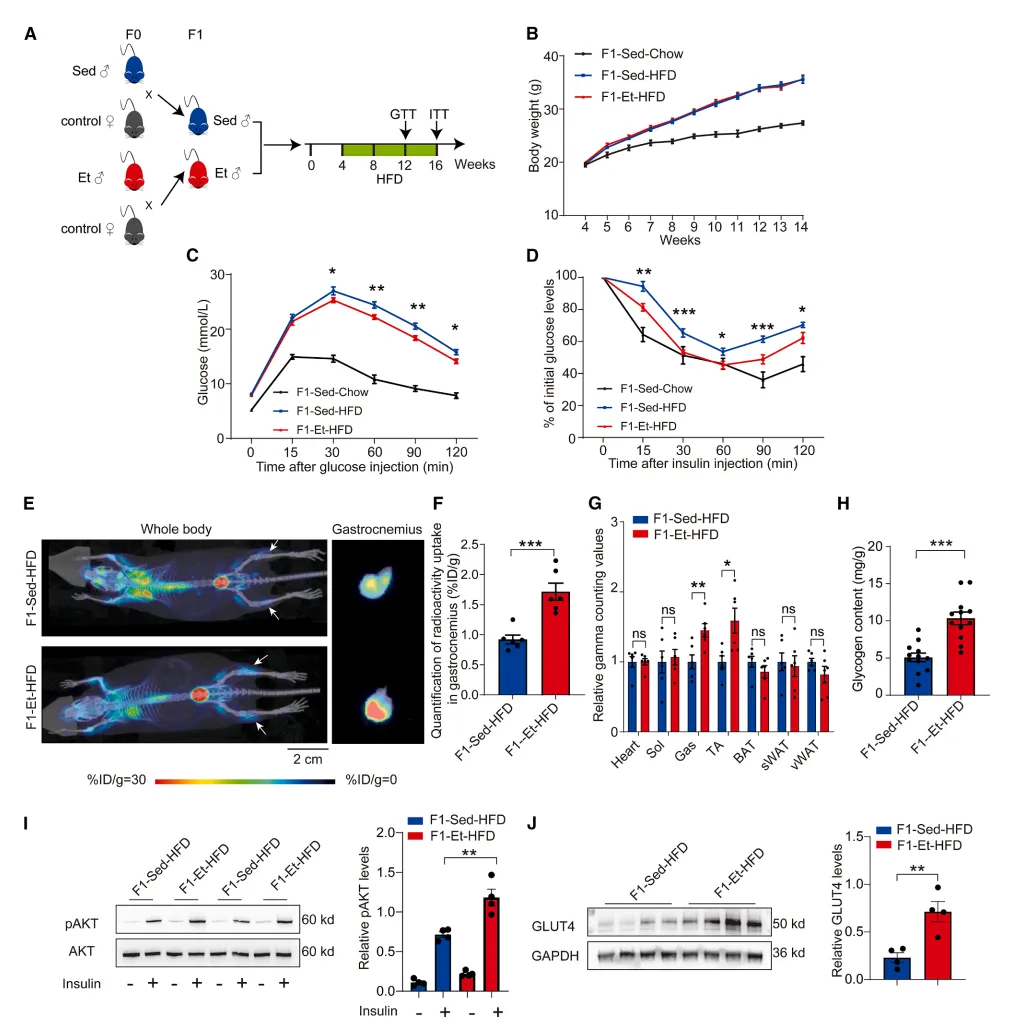
不止提升运动耐力,父代运动还留给孩子一层代谢保护buff:子代长期吃高脂高热量饮食时,更不容易发胖、不容易出现胰岛素抵抗、血糖紊乱。借助PET/CT精准追踪发现,这份保护不是全身泛化作用,而是特异性强化骨骼肌的葡萄糖摄取、糖原储存和胰岛素敏感性,从根源降低后代成年后代谢综合征风险【图3】。
图3.父代运动可提升子一代后代的葡萄糖摄取能力与胰岛素敏感性
小编总结:父代孕前运动相当于给孩子提前接种 “代谢防护 buff”,长远降低肥胖和糖尿病易感风险。
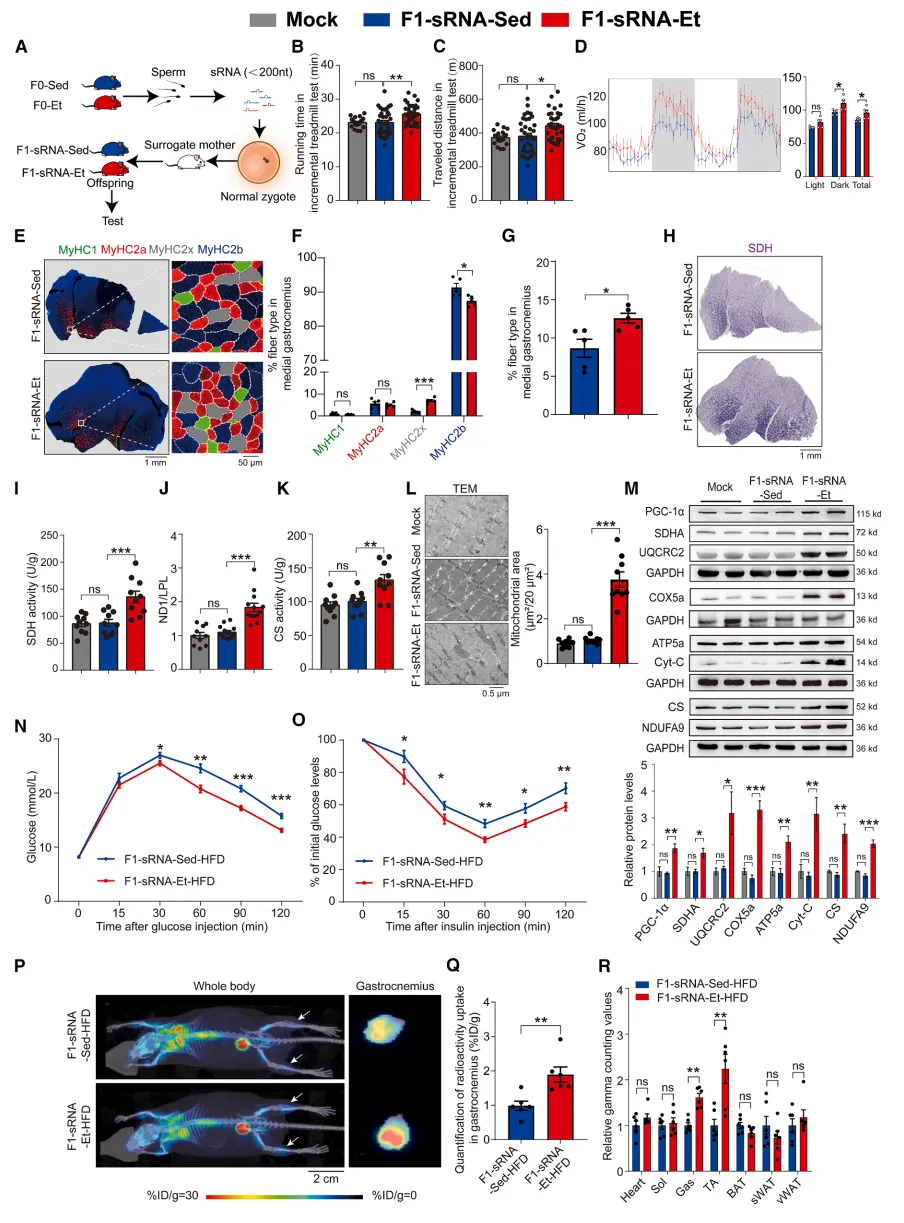
功能性验证精子小RNA是传递运动获益的充分必要介质
研究做了经典的功能回补实验:把运动父本精子的总RNA、小RNA、长RNA分别注入正常受精卵,只有精子小RNA能完整复刻子代高耐力、高代谢、线粒体优化的所有表型。直白说:父亲后天运动形成的生理优势,不靠DNA序列改变,而是打包藏在精子小RNA里,受精时直接交给胚胎、改写子代发育程序【图4】。
图4.精子小核糖核酸可将运动诱导的生理及代谢适应性特征,通过父代遗传给子一代后代
小编总结:正式实锤精子小RNA是承载父代运动记忆、实现跨代表观遗传的核心载体。
鉴定运动特异性精子miRNA并发现其可抑制胚胎NCoR1
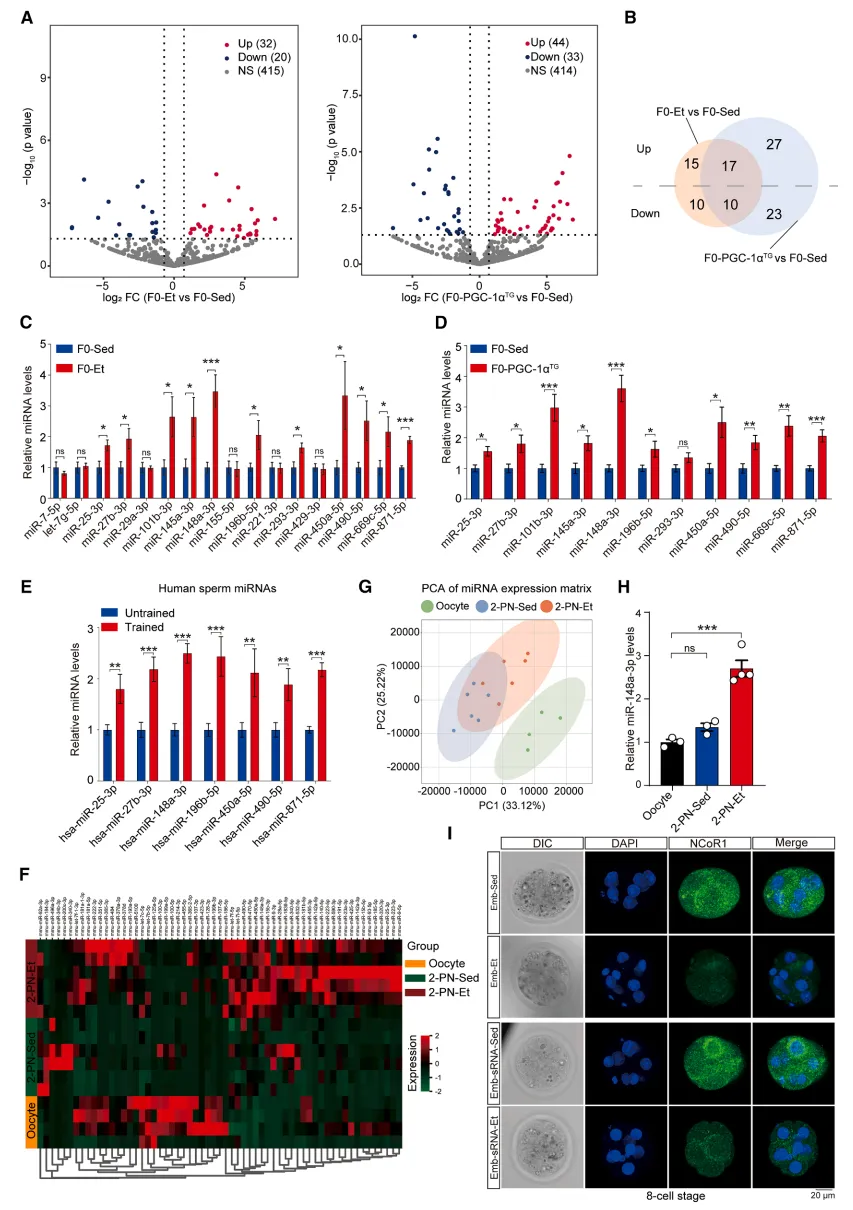
通过高通量测序对比,研究筛选出一批只在运动/高代谢父本精子中特异性上调的miRNA,且其中部分在人类运动男性精子中同样保守存在,提示这套机制在人体同样适用。这些特殊miRNA在受精后顺利进入早期胚胎,直接下调NCoR1蛋白表达。NCoR1本身是强转录抑制因子,相当于 “刹车”;父源miRNA把刹车松开,胚胎代谢发育通路被提前激活【图5】。
图5.父亲运动干预下精子微小RNA变化的评估及其向卵母细胞高效传递的验证
小编总结:锁定了运动改写的精子miRNA分子密码,打通了精子信号调控胚胎发育的关键节点。
确立胚胎NCoR1是精子miRNA行使跨代编程功能的关键靶点
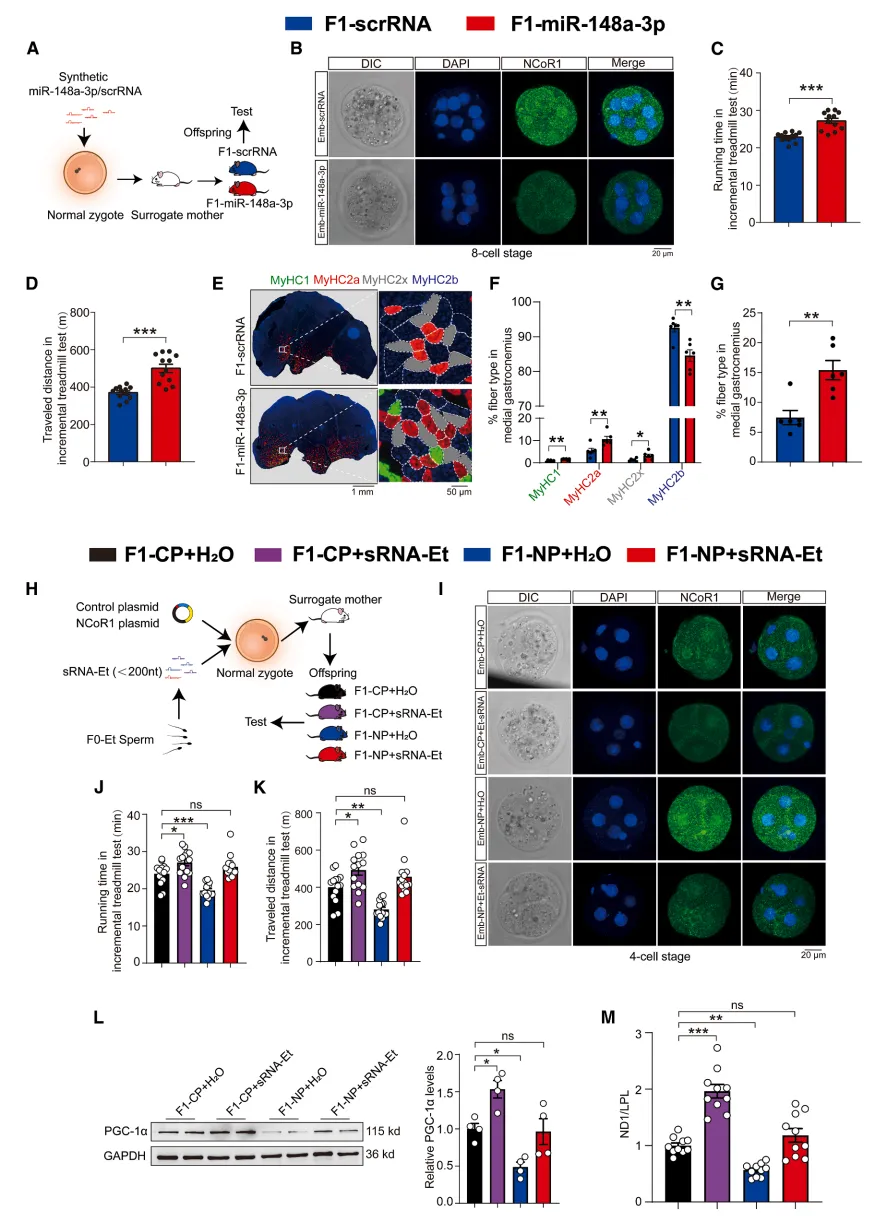
研究最后做了严谨的挽救实验:在胚胎中强行恢复NCoR1表达后,原本精子小RNA带来的高耐力、线粒体优化、代谢获益全部被抵消。这就彻底坐实整条通路:父代运动→精子miRNA重塑→胚胎抑制NCoR1→解除PGC-1α通路抑制→子代骨骼肌线粒体与耐力先天升级【图6】。
图6.将miR-148a-3p注入受精卵可提升子代的耐力,而在早期胚胎中恢复 NCoR1的表达则会削弱该效果
小编总结:通过反向挽救实验完成因果闭环,彻底阐明了整条信号通路的调控逻辑。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
