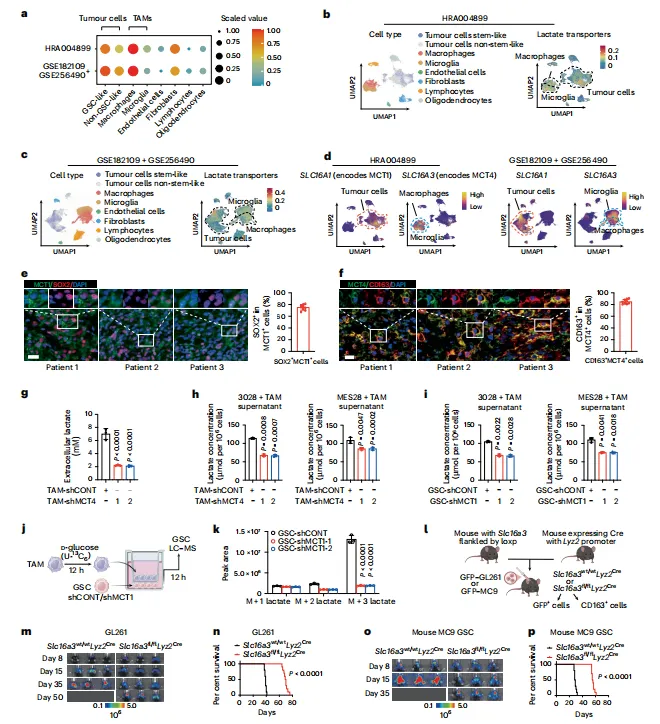
单细胞测序显示,SLC16A3(MCT4)在TAM高表达,SLC16A1(MCT1)在GSC富集。代谢流验证了乳酸从巨噬细胞流向肿瘤细胞。这一步空间定位做得很干净,细胞互作逻辑一下就有了(图1)。
图1:胰腺导管癌干细胞与肿瘤相关巨噬细胞中的乳酸代谢及转运蛋白特性
单细胞定位做得很扎实,但样本量仅7例IDH野生型患者,肿瘤异质性可能被低估。建议扩大队列验证MCT1/MCT4共表达模式是否普遍存在。
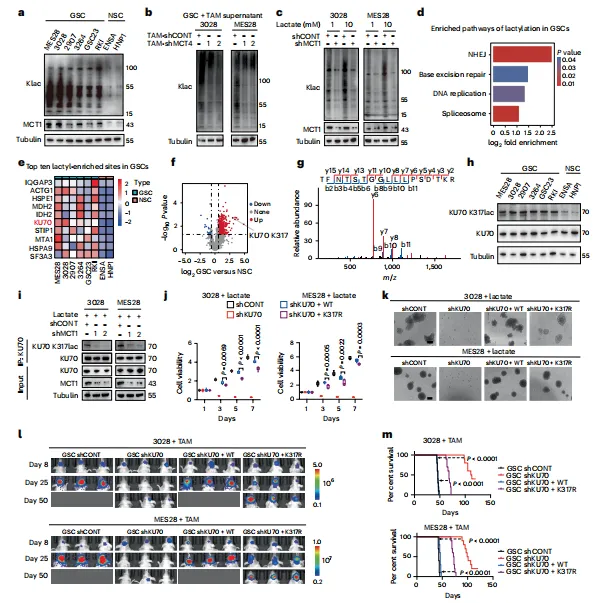
敲低TAM的MCT4,上清乳酸降了70%;敲低GSC的MCT1,13C标记乳酸摄入显著减少。功能互补实验确认了该转运轴对GSC增殖是必需的。这种双向验证设计,说服力直接拉满(图2)。
图2:通过MCT1–MCT4介导的乳酸在胶质干细胞与肿瘤相关巨噬细胞之间的转运促进GSCs增殖
代谢流示踪设计严谨,双向敲低互为验证。风险在于体外共培养无法完全模拟体内TAM与GSC的空间毗邻关系,原位验证会更稳妥。
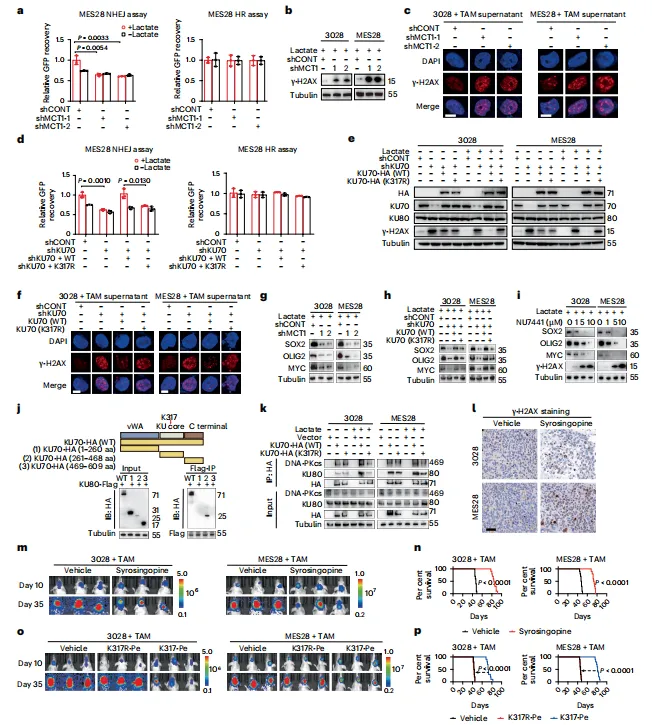
质谱筛选出GSC特异的KU70 K317乳酸化位点。回补实验显示,K317R突变体无法恢复增殖与成球能力。这说明该修饰位点比我们预想的更具功能权重,不只是被动副产物(图3)。
图3:TAM来源的乳酸可诱导DNA损伤修复蛋白KU70 K317发生乳酰化,从而导致生殖干细胞增殖
质谱筛选到新位点值得肯定,但K317R回补仅部分恢复表型,说明可能存在其他协同修饰位点未被捕获。建议同步检查乙酰化交叉调控。
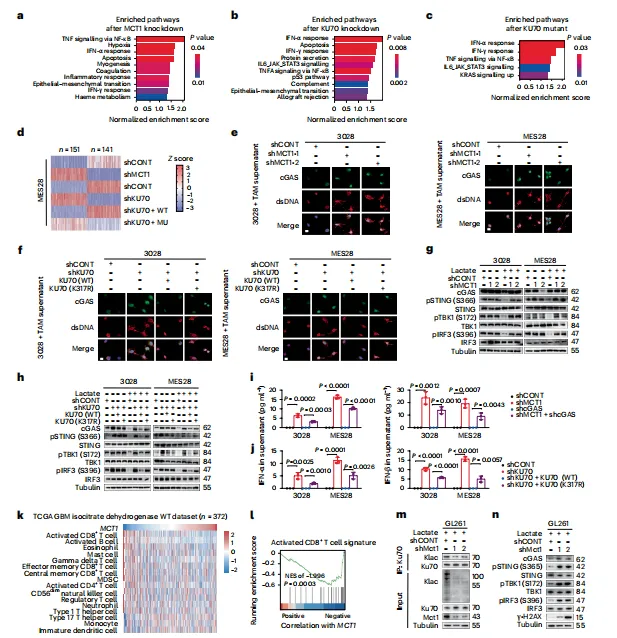
MCT1或KU70缺失增强γ-H2AX信号,NHEJ报告系统证实修复效率受损。乳酸化缺失导致KU70-KU80结合减弱。看来乳酸化直接调控了修复复合物的组装速度,这一点值得后续深挖(图4)。
图4:KU70 K317位点的乳酸化修饰可增强 NHEJ 功能并维持生殖干细胞的干性特征
NHEJ报告系统区分了HR与NHEJ贡献,这点设计巧妙。风险是KU70-KU80结合实验仅用免疫沉淀,缺乏结构生物学支撑,结合亲和力变化无法定量。
KU70 K317乳酸化减少了胞质dsDNA积累与cGAS-STING活化。下游IFN-α/β分泌下降,CD8+T细胞功能受抑。这条通路串联很完整,把代谢修饰与免疫逃逸直接挂上了钩(图5)。
图5:KU70 K317位点的乳酰化可抑制cGAS–sting信号通路并维持免疫抑制 TME
cGAS-STING这条线串联得很完整,但胞质dsDNA的来源未明确排除线粒体DNA泄漏。加DNase处理和线粒体膜电位检测会更严谨。
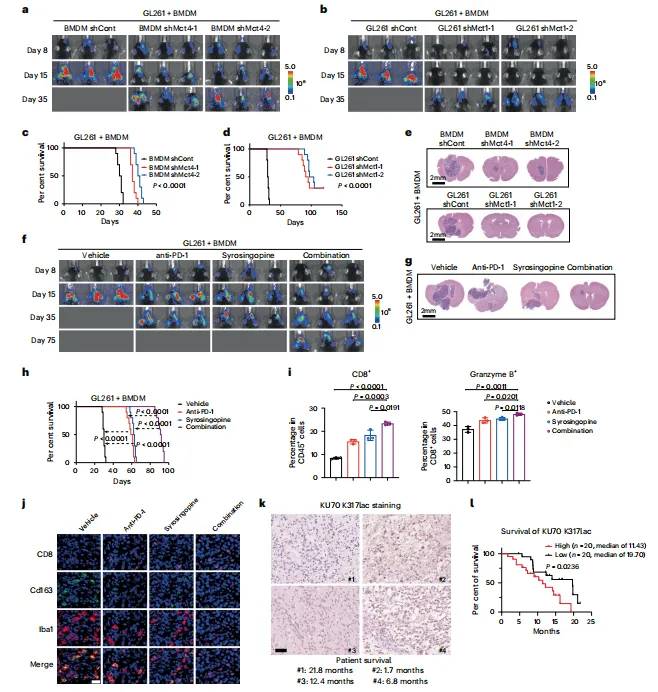
MCT抑制剂syrosingopine联用抗PD-1,肿瘤负荷降低最显著,CD8+T细胞浸润增加。在免疫完整模型中,组合策略优于单药。这为临床试验提供了一个可操作的配伍方案(图6)。
图6:乳酸转运体抑制剂联合PD-1抗体治疗 GBM 的疗效
联合治疗数据漂亮,但syrosingopine对MCT1/4的选择性不算高,可能带来脱靶效应。后续用条件敲除小鼠做治疗验证会更干净。







 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
