研究背景与目的
猪丁型冠状病毒(PDCoV)是近年来新流行的肠道冠状病毒,可感染各年龄阶段猪,引起仔猪腹泻、呕吐、脱水甚至死亡,临床症状与PEDV、TGEV相似,且常混合感染,给猪肠道腹泻病防控带来巨大困难。PDCoV具有跨物种传播潜力,2021年《Nature》报道在海地儿童血浆中分离出PDCoV毒株,提示其人畜共患风险。冠状病毒辅助蛋白虽不作为病毒粒子主要成分,但对病毒吸附、增殖及逃避宿主天然免疫至关重要。PDCoV辅助蛋白NS6已被证实通过干扰RIG-I/MDA5与dsRNA结合抑制干扰素诱导。NS7蛋白的功能尚不明确,其内部NS7a为干扰素拮抗剂,可通过破坏IKKε与TRAF3和IRF3结合抑制IFNβ产生。本研究旨在利用本实验室构建的基于BAC系统的PDCoV反向遗传平台,通过CRISPR-Cas9技术构建NS7基因缺失重组病毒(rPDCoV-ΔNS7),系统评估NS7缺失对PDCoV体外复制特性及仔猪体内致病力的影响,为PDCoV致病机制研究及疫苗靶点筛选提供理论依据。
1 材料与方法
1.1 毒株、细胞、质粒及试验动物
毒株: PDCoV JS2021-LX毒株(GenBank: OQ473581.1);细胞与质粒: LLC-PK1细胞;pBAC-PDCoV反向遗传操作系统。试验动物: PEDV、TGEV、PoRV、PDCoV抗原抗体阴性仔猪。
1.2 主要试剂
抗体: PDCoV NS7蛋白多克隆抗体;PDCoV N蛋白单克隆抗体;CoraLite 594标记羊抗鼠IgG;羊抗鼠IgG-HRP。试剂盒与工具酶: Lipofectamine 3000(Thermo);CRISPR-Cas9内切酶、T7体外转录试剂盒(NEB);RNA提取试剂盒(Omega);RNA反转录试剂盒、同源重组试剂盒、DNA Marker(诺唯赞);质粒大提试剂盒(MACHEREY NAGEL);DH10B感受态细胞(昂羽生物)
1.3 pBAC-PDCoV-ΔNS7重组质粒的构建
策略设计: NS7基因嵌套于N基因开放阅读框内,采用同义突变策略——将NS7基因内所有起始密码子ATG突变为ACG(苏氨酸),实现NS7蛋白功能选择性失活且不改变N基因编码氨基酸序列。
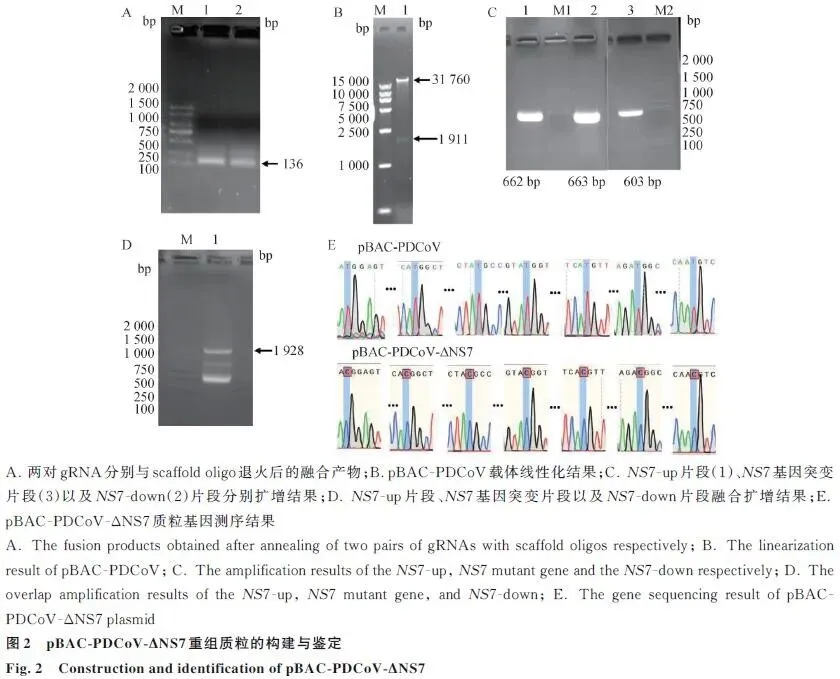
技术路线:gRNA设计:在NS7基因上下游各设计一对gRNA,与scaffold oligo退火形成双链,T7体外转录获得sgRNAΔNS7a和sgRNAΔNS7b;Cas9切割:Cas9内切酶与sgRNA定位切割pBAC-PDCoV载体,去除NS7基因,获得线性化载体;片段扩增:分别扩增NS7-up片段(662 bp)、NS7-down片段(663 bp)及NS7基因突变片段(603 bp);同源重组:通过overlap PCR融合三个片段,与线性化载体同源重组连接;转化验证:连接产物转化DH10B感受态细胞,单克隆测序验证。
1.4 rPDCoV-ΔNS7病毒的拯救与鉴定
病毒拯救: pBAC-PDCoV-ΔNS7重组质粒转染单层LLC-PK1细胞,10h后换含10 μg·mL⁻¹胰蛋白酶的DMEM维持液培养,待细胞病变(CPE)后冻融离心收集上清(F1代),传代三次。
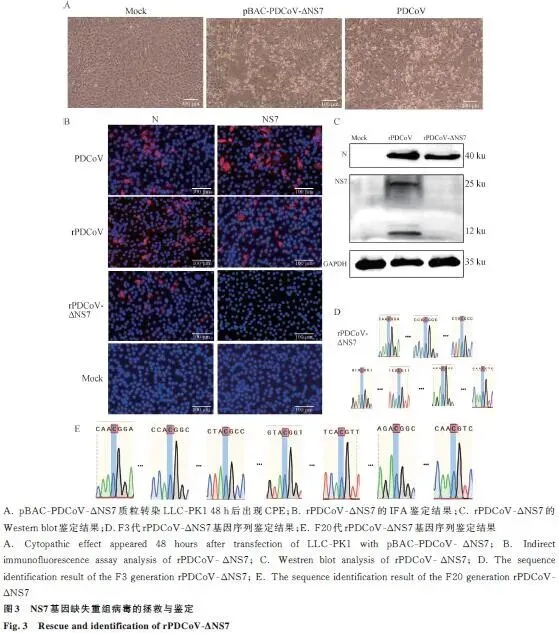
鉴定方法:间接免疫荧光(IFA): 检测N蛋白和NS7蛋白表达;Western blot: 验证NS7蛋白缺失及N蛋白表达;RT-PCR测序: F3代验证NS7基因突变,F20代验证遗传稳定性。
1.5 PDCoV、rPDCoV以及rPDCoV-ΔNS7的体外生物学特性比较
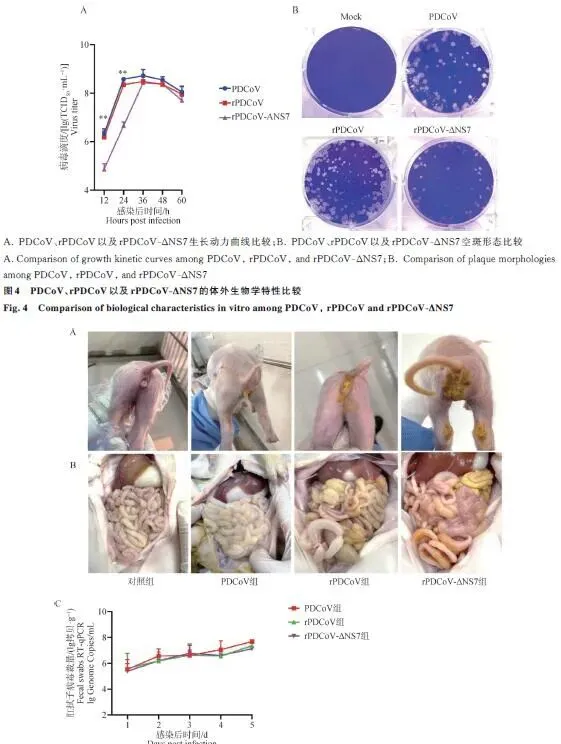
生长动力学: 三种病毒均以0.01 MOI接种LLC-PK1细胞,吸附2h后换含8 μg·mL⁻¹胰蛋白酶的维持液,分别于12、24、36、48、60h收取病毒上清测定TCID₅₀,绘制生长曲线。
空斑形态: 同样剂量接种LLC-PK1细胞,2×DMEM与1.8%低熔点琼脂糖1:1覆盖,37 °C培养48 h,结晶紫染色观察空斑大小。
1.6 仔猪致病性试验
分组设计: 4日龄仔猪12头,分4组(对照组、PDCoV组、rPDCoV组、rPDCoV-ΔNS7组),每组3头,口服攻毒,剂量10⁷ TCID₅₀·头⁻¹。
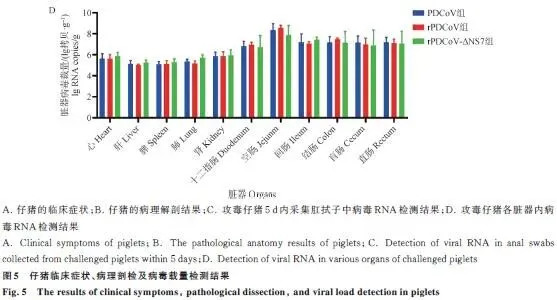
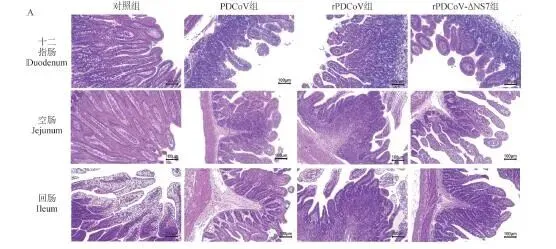
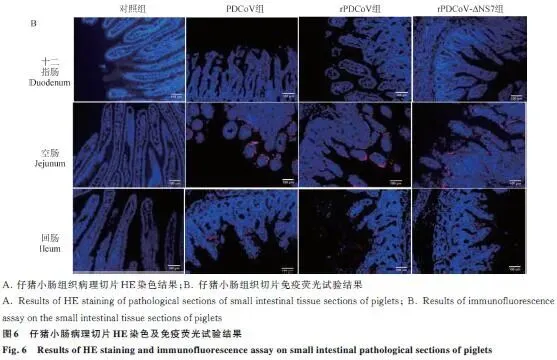
监测指标:每日观察精神状态、食欲、腹泻症状;攻毒24 h后每日采集肛拭子,RT-qPCR检测病毒载量;濒死仔猪立即剖杀,存活仔猪攻毒后第5天统一剖杀;采集心、肝、脾、肺、肾及肠道各段,制备组织悬液检测病毒分布;十二指肠、空肠、回肠固定,HE染色和免疫荧光分析病理变化。
1.7 数据分析
GraphPad 8.0统计分析,one-way ANOVA或two-way ANOVA比较组间差异,P<0.05为显著。
2 结果
2.1 pBAC-PDCoV-ΔNS7重组质粒的构建与鉴定
gRNA退火产物: 两对gRNA与scaffold oligo退火融合产物大小136 bp,电泳验证正确。载体线性化: pBAC-PDCoV经Cas9酶切后可见31 760和1 911 bp两条带,切割成功。片段扩增: NS7-up(662 bp)、NS7突变片段(603 bp)、NS7-down(663 bp)及overlap融合产物(1 928 bp)大小均符合预期。测序验证: 单克隆测序显示原NS7基因被成功替换为突变序列,ATG→ACG突变位点准确无误(图2)。
2.2 rPDCoV-ΔNS7的拯救与鉴定
CPE观察: 质粒转染LLC-PK1细胞48 h后出现典型细胞病变。IFA鉴定: rPDCoV-ΔNS7感染组可检测到N蛋白(红色荧光),检测不到NS7蛋白;PDCoV和rPDCoV组N蛋白和NS7蛋白均阳性。Western blot: rPDCoV-ΔNS7组N蛋白表达正常,NS7蛋白缺失;GAPDH内参一致。遗传稳定性: F3代和F20代测序均显示NS7基因突变位点稳定存在,无其他突变发生。结论: 基于BAC系统的NS7基因缺失重组病毒成功构建且遗传稳定(图3)。
2.3 PDCoV、rPDCoV以及rPDCoV-ΔNS7的体外生物学特性比较
生长动力学: 三种病毒生长曲线均呈先升后降趋势,36 h达复制峰值(均超10⁸ TCID₅₀·mL⁻¹)。rPDCoV-ΔNS7在12和24 h时病毒滴度显著低于PDCoV和rPDCoV(P<0.01),但36h达到峰值后与两组无统计学差异(P>0.05)。空斑形态: 三种病毒在LLC-PK1上形成的空斑大小无明显差异。分析: NS7缺失导致病毒早期复制延迟,但后期可补偿至正常水平,提示NS7可能参与早期免疫逃逸而非病毒复制必需(图4)。
2.4 仔猪致病性试验分析
临床症状: 三组攻毒仔猪24 h后均出现精神沉郁、食欲下降,40 h左右个别出现腹泻,96 h症状减轻;对照组正常。三组临床症状无差异。病理剖检: 三组仔猪均见小肠胀气、肠壁变薄,以空肠病变最明显,组间无差异。排毒监测: 肛拭子RT-qPCR显示三组仔猪排毒水平无统计学差异(P>0.05),24 h可检出病毒,随感染时间延长载量增加。组织嗜性: 心、肝、脾、肺、肾及各段肠道均能检测到病毒,空肠病毒载量最高,三组组织内病毒分布无显著差异。病理切片:HE染色: 三组仔猪十二指肠、空肠、回肠均出现肠绒毛变短、萎缩甚至断裂,与对照组差异明显,组间无差异。免疫荧光: 病毒抗原信号主要分布在空肠,回肠次之,十二指肠最少,三组荧光信号强度无明显差异(图5、6)。
3 讨论
反向遗传系统比较: BAC系统在大肠杆菌中操作,避免大量活病毒体外操作,降低实验室感染风险,基因编辑更精准,拯救病毒更接近自然感染状态。与Zhang等(2021)体外连接转录RNA拯救方法相比,本研究BAC系统拯救的rPDCoV-ΔNS7在12和24 h病毒滴度显著降低,而Zhang等报道NS7缺失不影响增殖效果。差异可能源于:①BAC系统通过细菌繁殖保证全长基因组完整性;②体外转录RNA可能引发更强炎症反应,激活宿主修复机制部分补偿基因缺失影响。
NS7功能定位: NS7a作为干扰素拮抗剂,通过破坏IKKε-TRAF3-IRF3复合物抑制IFNβ产生。rPDCoV-ΔNS7早期复制延迟可能与NS7蛋白免疫调控功能缺失相关,宿主早期先天免疫应答增强抑制病毒复制;36 h后病毒滴度恢复,提示存在其他免疫逃逸机制补偿或宿主免疫应答衰减。
疫苗靶点评估: NS7基因缺失不影响病毒最终复制水平和致病性,表明NS7不是PDCoV复制的必需基因,也非重要毒力基因,不适合作为基因缺失弱毒疫苗的靶基因。但NS7对宿主免疫应答的调控效应尚需通过系统性免疫试验进一步阐明。
研究展望: 综合评估NS7缺失重组病毒对宿主天然免疫及适应性免疫应答的影响,有助于明确NS7在病毒免疫逃逸中的潜在功能,为PDCoV防控策略优化提供新思路。
4 结论
1) 基于BAC-CRISPR/Cas9系统成功拯救PDCoV JS2021-LX毒株NS7基因缺失重组病毒(rPDCoV-ΔNS7),遗传稳定性良好(F20代验证)。2) NS7缺失导致病毒早期复制延迟(12、24 h滴度显著降低),但36 h后恢复至正常水平,空斑形态无差异。3) NS7缺失不影响仔猪临床症状、病理损伤、排毒规律及组织嗜性,与亲本病毒和野生型拯救病毒无显著差异。4) NS7不是PDCoV复制必需基因和重要毒力基因,不适合作为弱毒疫苗靶点;但其免疫调控功能值得深入研究。
关键词:PDCoV ; 感染性克隆 ; 基因缺失 ; 攻毒试验 ; 反向遗传













 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
