DOI:10.1016/j.xplc.2026.101973
植物在抵御病原菌侵染时需要迅速积累水杨酸(Salicylic acid,SA),但过量或持续积累的SA会加速叶片衰老。那么,植物如何在完成防御后及时终止SA合成,长久以来尚不清晰。近日,南京农业大学生命科学学院常明团队联合河南农业大学王燃团队阐明E3泛素连接酶PUB13通过靶向降解SA生物合成关键酶PBS3,促进病原菌诱导SA积累的适时回落,从而协调植物免疫与叶片寿命平衡。相关研究成果以“PUB13-Mediated Degradation of PBS3 Regulates Salicylic Acid Biosynthesis to Coordinate Plant Immunity and Leaf Longevity”为题发表于 Plant Communications。
水杨酸(Salicylic acid,SA)是植物免疫系统中最重要的激素之一。当植物遭受病原菌侵染时,体内SA迅速积累,激活局部防御反应和系统获得性抗性(Systemic Acquired Resistance,SAR),从而提高对病原菌的抵抗能力。然而,免疫反应并非越强越好。长期维持高水平SA虽然能够增强抗病性,却会抑制生长并加速叶片衰老。因此,植物需要一套精准的调控机制:既要在病原菌侵染时迅速启动SA生物合成,又必须在防御建立后及时关闭这一过程,以避免长期免疫激活带来的生长代价。PBS3(AVRPPHB SUSCEPTIBLE 3,也称GH3.12)是十字花科植物拟南芥SA生物合成途径中的关键酶,被认为是控制SA产生的重要“阀门”。然而,PBS3蛋白如何被动态调控,以及植物如何在免疫激活后终止SA持续积累,长期以来并不清楚。
围绕这一科学问题,研究团队综合运用遗传学、生物化学和分子生物学等研究手段,系统解析了PBS3蛋白稳定性的调控机制。研究发现,PBS3含量直接决定植物在抗病与衰老之间的平衡。PBS3缺失突变体表现为SA积累不足、抗病能力下降,但叶片衰老明显延缓;相反,随着PBS3表达量逐渐升高,病原菌诱导的SA积累、系统获得性抗性以及叶片衰老程度均同步增强,表明PBS3是协调植物免疫与叶片寿命的重要决定因子。
为了寻找PBS3的上游调控因子,研究人员通过酵母双杂交(Y2H)筛选获得E3泛素连接酶PUB13。进一步利用双分子荧光互补(BiFC)、GST pull-down以及免疫共沉淀(Co-IP)等实验,证实PUB13能够直接与PBS3结合,并介导PBS3发生泛素化修饰,最终通过26S蛋白酶体途径将其降解。
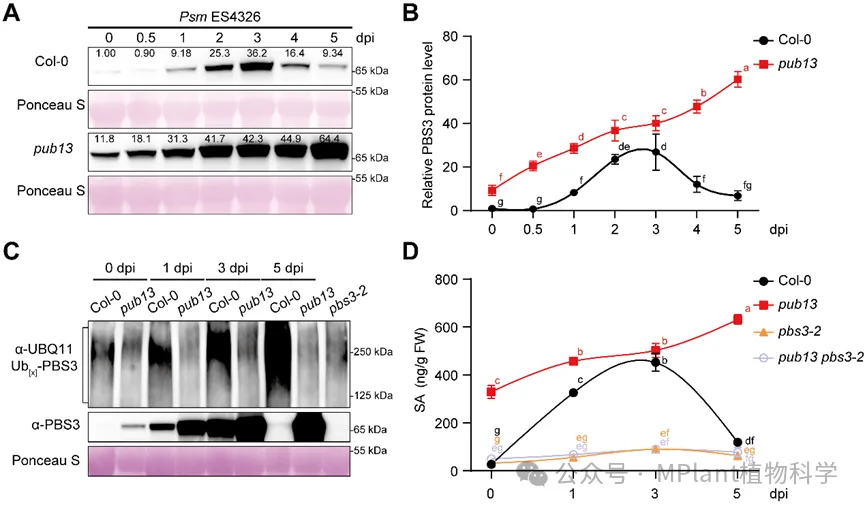
进一步研究发现,在病原菌侵染后,野生型拟南芥Col-0中的PBS3蛋白和SA含量均呈现“先升高、后下降”的动态变化模式(图1)。PBS3蛋白在病原菌侵染后迅速积累,驱动SA大量合成;随后PUB13介导PBS3降解,使SA水平逐渐回落,从而终止过度的免疫反应。而在pub13突变体中,这一“关闭开关”失效(图1)。PBS3无法被有效降解,导致SA在达到峰值后仍持续维持高水平状态。虽然植物获得了更强的系统抗性,但同时出现明显加速的叶片衰老现象。
图1 PUB13促进病原菌诱导水杨酸积累的及时消退
遗传学分析进一步表明,PBS3位于PUB13下游发挥作用。缺失PBS3能够完全抑制pub13突变体中增强的抗病性和早衰表型,说明PUB13主要通过调控PBS3稳定性来影响SA生物合成及其下游生理过程。
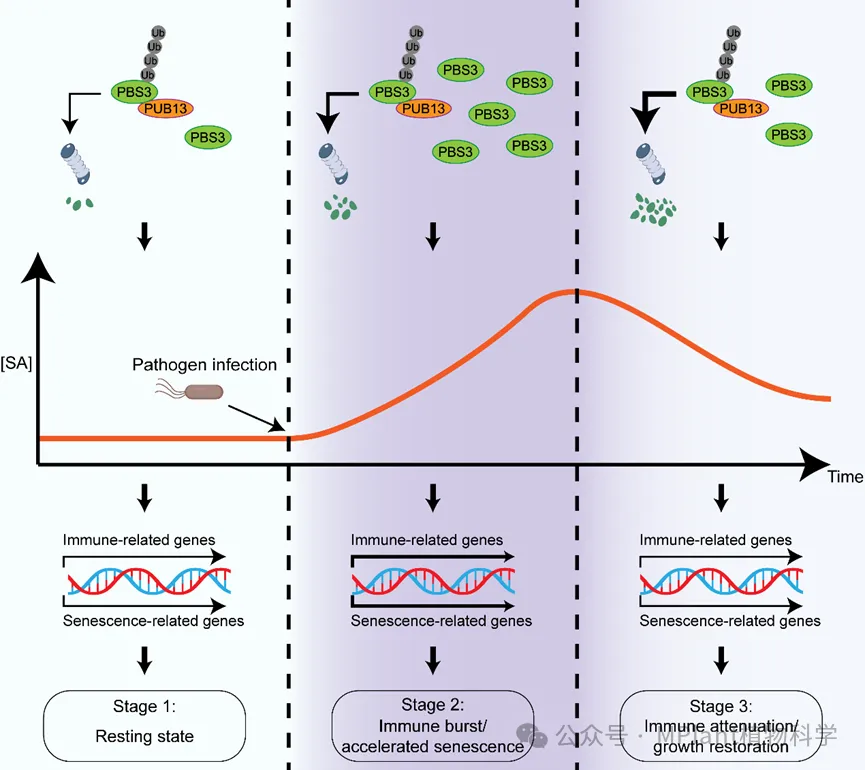
综合遗传学、生化和表型分析结果,研究团队构建了PUB13–PBS3调控植物免疫与叶片寿命平衡的工作模型(图2)。
图2 PUB13–PBS3模块调控植物免疫与叶片寿命平衡的工作模型
病原菌侵染后,PBS3迅速积累并促进SA生物合成,激活植物免疫反应和系统获得性抗性;随着防御建立,PUB13介导PBS3泛素化降解,通过26S蛋白酶体途径降低PBS3蛋白水平,促进SA积累及时消退,使植物逐步恢复生理稳态。在pub13突变体中,由于PBS3不能被有效降解,病原菌诱导的SA积累长期维持高水平,虽然增强了植物抗病能力,但同时导致叶片过早衰老。
该研究明确了PUB13作为SA合成的负向调控开关,通过动态调控PBS3蛋白周转限制SA过量积累,完善了植物免疫稳态调控理论,同时为培育兼具强抗病、正常生长特性的作物提供全新分子靶点。
南京农业大学生命科学学院常明教授和河南农业大学王燃教授为论文共同通讯作者。南京农业大学博士研究生何金宇和李围博士为论文共同第一作者。该研究得到国家自然科学基金等项目资助。








 10个月宝宝每天需要喝多少奶粉?
10个月宝宝每天需要喝多少奶粉?
