南京农大发Science正刊,今年第三篇!
- 2026-07-17 04:17:51


2026年7月2日,Science在线发表了来自南京农业大学董莎萌教授团队和英国帝国理工学院 Tolga Bozkurt共同为通讯作者,题为“Surface immune signaling unlocks NLR activation through mRNA alternative splicing”的研究论文。该研究揭示了植物表面免疫信号通过促进NLR免疫受体基因Rpi-vnt1.1的mRNA可变剪切,移除其N端抑制性结构域,从而解锁胞内NLR免疫受体激活的崭新机制。这一发现阐明了植物模式触发免疫(PTI)与效应子触发免疫(ETI)两大防御系统之间的协同调控机制,为理解植物如何平衡免疫激活与自身免疫损伤提供了重要的分子基础。
值得注意的是,这是南京农业大学今年第三篇Science, 之前两篇Science分别为:南京农大发Science正刊,破解水稻“增根抑冠”之谜,南京农业大学发Science正刊
植物依赖细胞表面的模式识别受体(PRRs)和胞内核苷酸结合富亮氨酸重复序列(NLR)免疫受体分别识别病原微生物的保守分子特征和入侵的效应蛋白,激活PTI和ETI以抵御病害。尽管越来越多的证据表明PTI和ETI在功能上具有协同作用,但PTI究竟如何调控ETI的激活,其精确分子机制长期以来尚不明确。在大多数卷曲螺旋型NLR(CNL)中,其N端的MADA基序是形成抗病小体(resistosome)和激活免疫所必需的结构。然而,部分NLR(包括具有重要农业价值的马铃薯晚疫病抗性蛋白Rpi-vnt1.1)在其MADA基序上游存在一段功能未知的N端延伸序列,这暗示着可能存在一种尚未被发现的调控机制,即通过移除该抑制性延伸来调控NLR的激活,从而在病原威胁存在时迅速启动免疫,而在无病原时防止自身免疫。
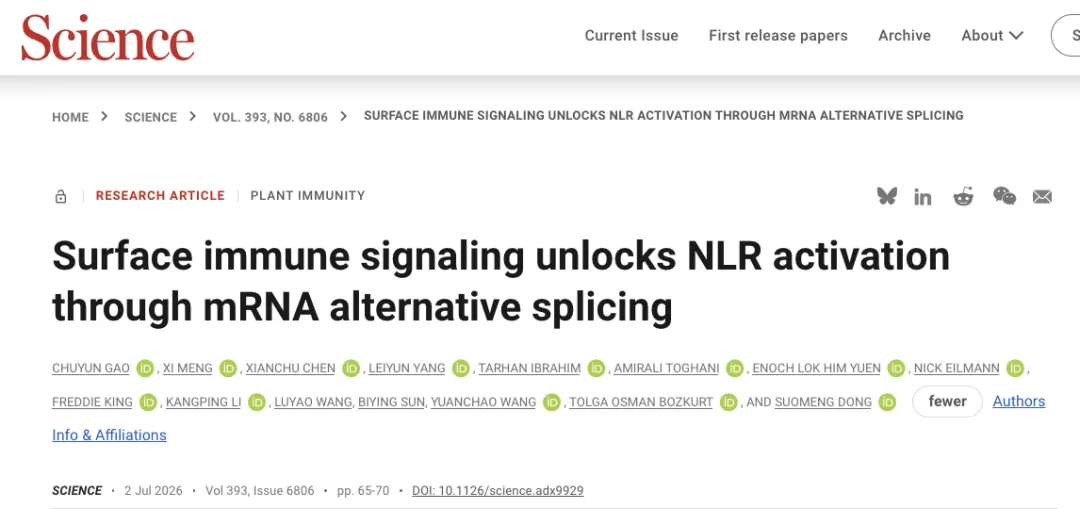
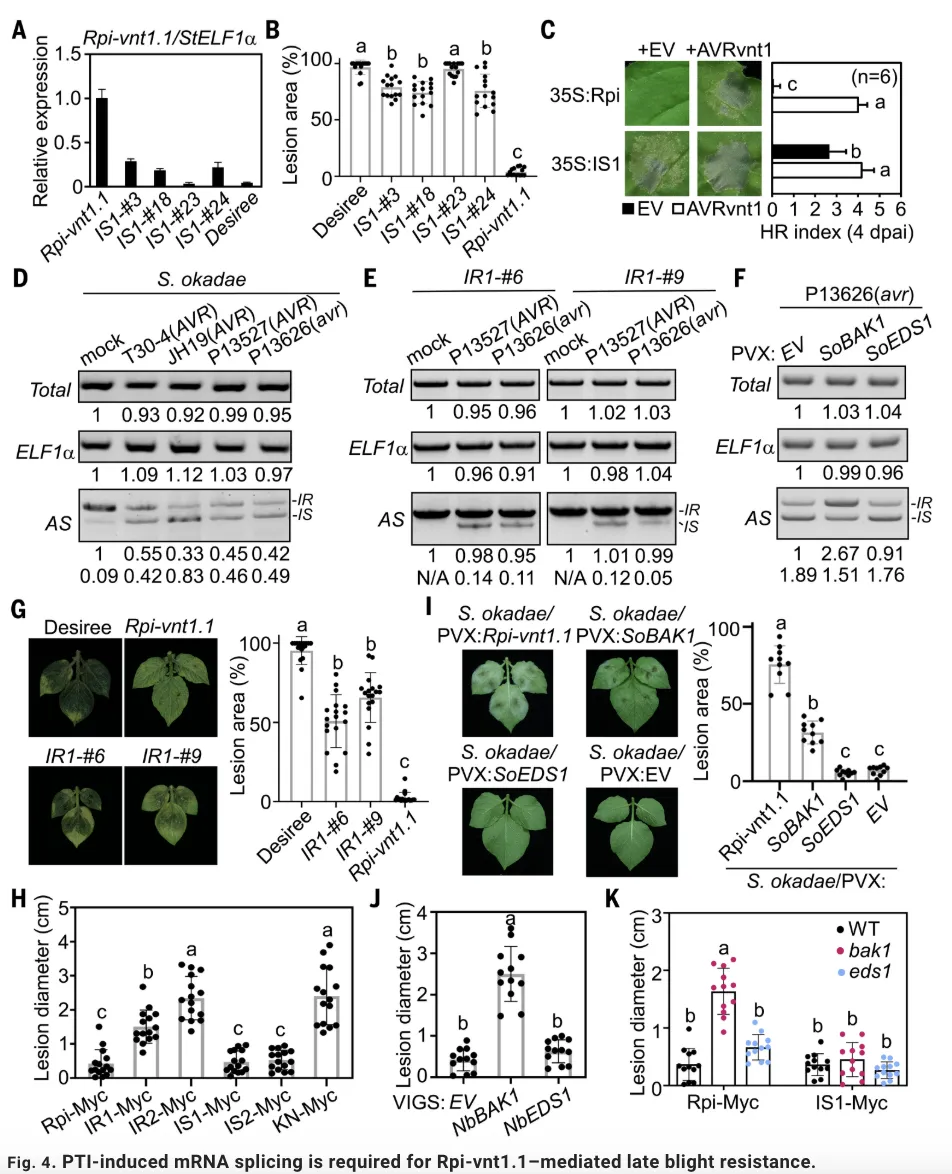
该研究首先通过一系列实验证实,PTI信号对于Rpi-vnt1.1介导的免疫反应完全激活是必需的。研究发现,用PAMPs(如INF1、flg22)处理表达Rpi-vnt1.1和其效应蛋白AVRvnt1的植物时,由Rpi-vnt1.1/AVRvnt1诱导的离子渗漏、过敏反应(HR)和过氧化氢积累均显著增强;而在PTI关键共受体BAK1的突变体中,Rpi-vnt1.1/AVRvnt1诱导的免疫反应则明显减弱。
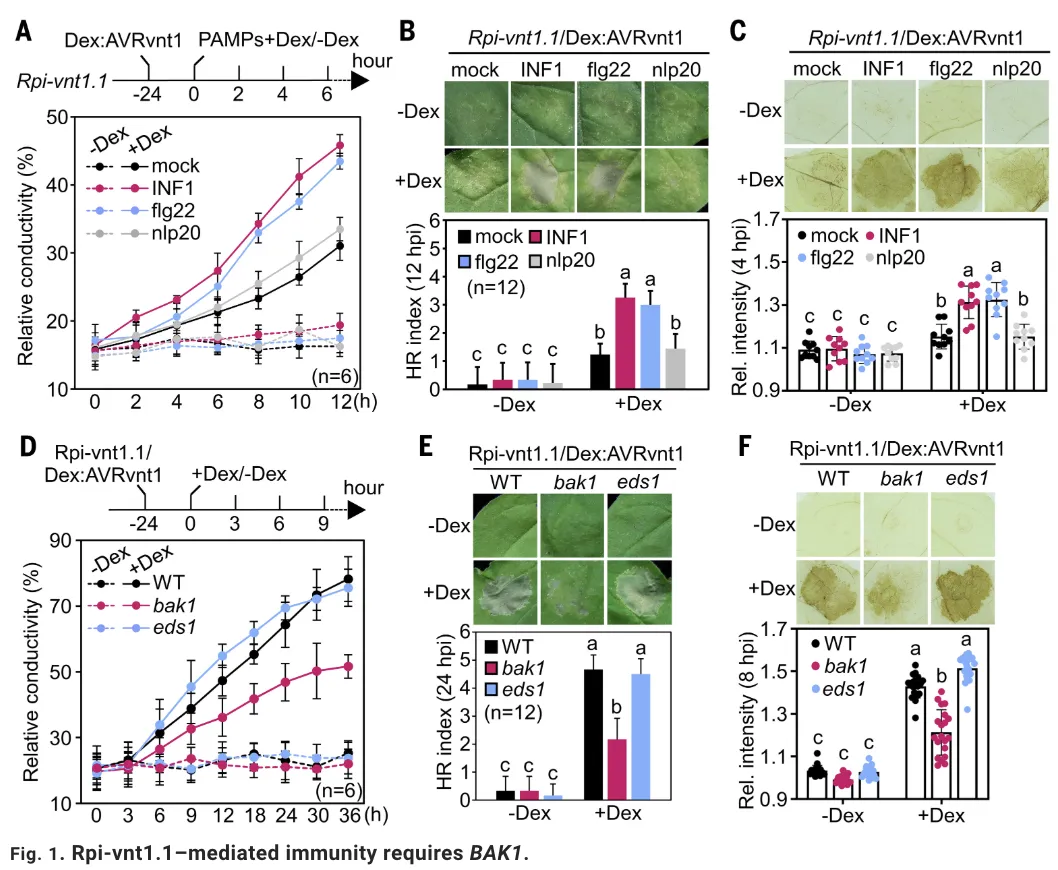
进一步的研究发现,PTI信号促进了Rpi-vnt1.1 mRNA的可变剪切。通过5′ RACE和转录组测序分析,研究团队鉴定出Rpi-vnt1.1基因5′区域存在一个U12型内含子,该内含子的保留(IR)或剪切(IS)产生两种不同的转录本异构体。PAMP处理(如flg22、INF1)能够显著促进该内含子的剪切,增加剪切异构体(Rpi-vnt1.1IS)的积累,而这一过程不依赖于nlp20处理,且不影响Rpi-vnt1.1总mRNA和蛋白水平。
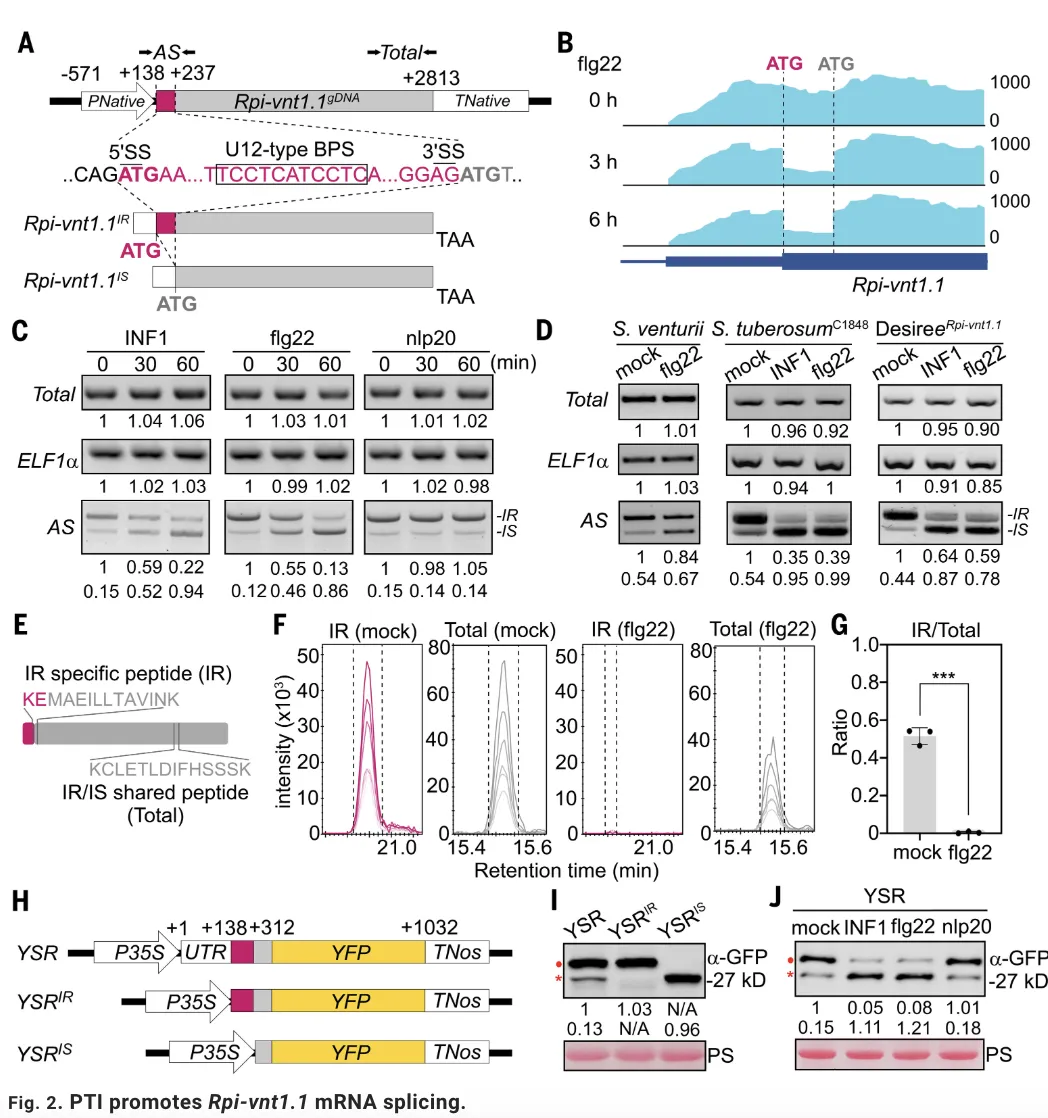
接下来,利用蛋白水平平行反应监测(PRM)质谱技术和构建YFP剪切报告系统,研究进一步证实PTI信号在蛋白水平上确实改变了Rpi-vnt1.1异构体的比例,使其向具有免疫活性的Rpi-vnt1.1IS变体倾斜。为明确剪切的功能意义,研究通过构建强制保留内含子或强制剪切的突变体,发现模拟剪切状态的Rpi-vnt1.1IS变体能够触发比野生型更强更快的免疫反应,表现为离子渗漏、HR和H2O2积累显著增加,而保留内含子的Rpi-vnt1.1IR变体则免疫活性大幅减弱甚至丧失。之后通过免疫共沉淀实验,研究证明只有剪切异构体Rpi-vnt1.1IS才能在效应蛋白AVRvnt1存在时形成强效的抗病小体寡聚化,而Rpi-vnt1.1IR则无法完成这一关键过程。
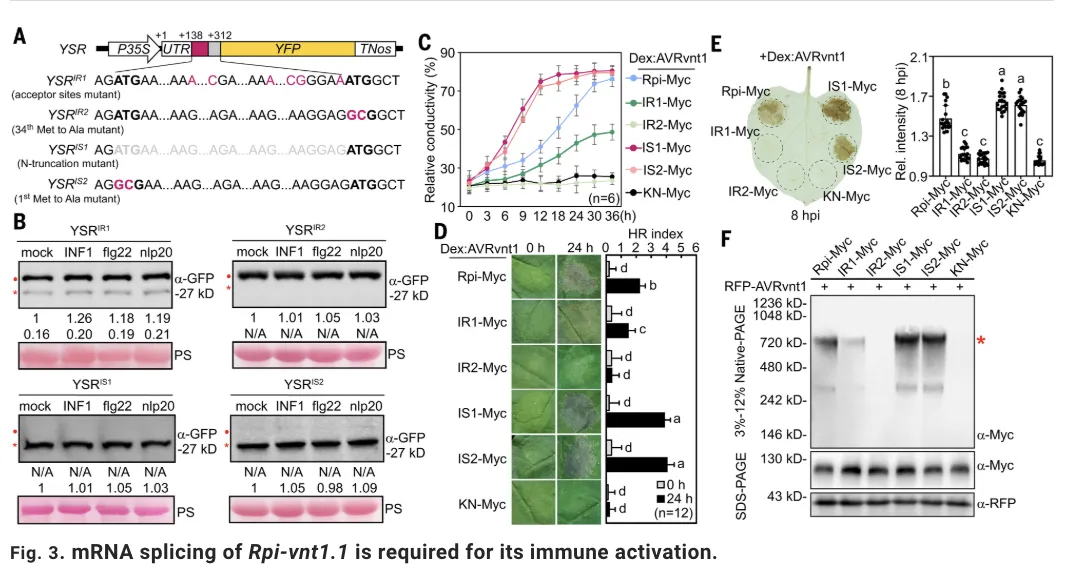
最终,在稳定转基因马铃薯和通过病毒诱导基因沉默(VIGS)的植株中进行的晚疫病抗性实验表明,PTI诱导的Rpi-vnt1.1 mRNA剪切是Rpi-vnt1.1介导的晚疫病抗性所必需的;在bak1突变体或BAK1沉默的植株中,Rpi-vnt1.1介导的抗性显著降低,而组成型表达的Rpi-vnt1.1IS变体所介导的抗性则不受影响,这确凿地证明了PTI通过促进mRNA剪切来增强Rpi-vnt1.1介导的ETI免疫。
该研究阐明了一种植物PTI信号通过调控NLR mRNA可变剪切来“解锁”ETI免疫激活的崭新分子机制,揭示了细胞表面受体信号如何翻译为胞内免疫受体的激活信号。这一发现不仅为理解植物两层免疫系统之间的协同联动提供了突破性的理论框架,还揭示了N端延伸作为一种广泛保守的“分子刹车”机制,在多种植物MADA-NLR中可能普遍存在,用于在静息状态下抑制NLR自身激活,同时保持对病原的灵敏响应。
论文链接:https://www.science.org/doi/10.1126/science.adx9929

