背景介绍
微塑料(MPs)已成为日益受到关注的新兴环境污染物,广泛分布于全球生态系统中。经口摄入被认为是人类暴露的主要途径——MPs已在多种日常食品和饮料中被检出,据估计成年人每周可能无意摄入约0.1–5克MPs。因此,胃肠道既是MPs暴露的首要部位,也是连接环境暴露与系统性健康效应的关键生物学界面。纳米塑料(NPs)因其更小的尺寸更易被吸收并在体内持久滞留。流行病学与实验证据提示MPs暴露与炎症性肠病之间存在强关联,且近年研究开始关注MPs对肠神经系统(ENS)的潜在影响,但NPs在肠道蓄积的驱动机制及其对ENS毒性的贡献尚不明确。
研究思路
针对上述问题,南京医科大学王婷教授、南京大学张徐祥教授团队合作,揭示了聚苯乙烯纳米塑料(PS-NPs)通过一条此前被忽视的“肝-胆-肠”肠肝循环通路显著增强其肠道滞留。通过口服暴露和尾静脉注射联合“Zombie”小鼠模型,研究证实被吸收的PS-NPs被肝脏高效捕获、浓缩于胆囊,随后经胆汁重新排入肠道。长期PS-NPs暴露导致明显的肠上皮损伤(杯状细胞丢失、紧密连接破坏)和细胞因子介导的炎症。多组学分析揭示肠道菌群失调、代谢物谱广泛改变及神经活性信号通路富集。机制上,PS-NPs通过下调抑制性神经递质VIP、上调TH和降低机械敏感通道PIEZO1表达诱导ENS损伤,进而损害肠道蠕动,形成“损伤—功能障碍—排泄延迟—再暴露”的自我放大循环。相关内容以Enterohepatic Circulation of Polystyrene Nanoplastics Promotes Intestinal Inflammation by Impairing Enteric Neurons发表在ACS Nano!
图片解析
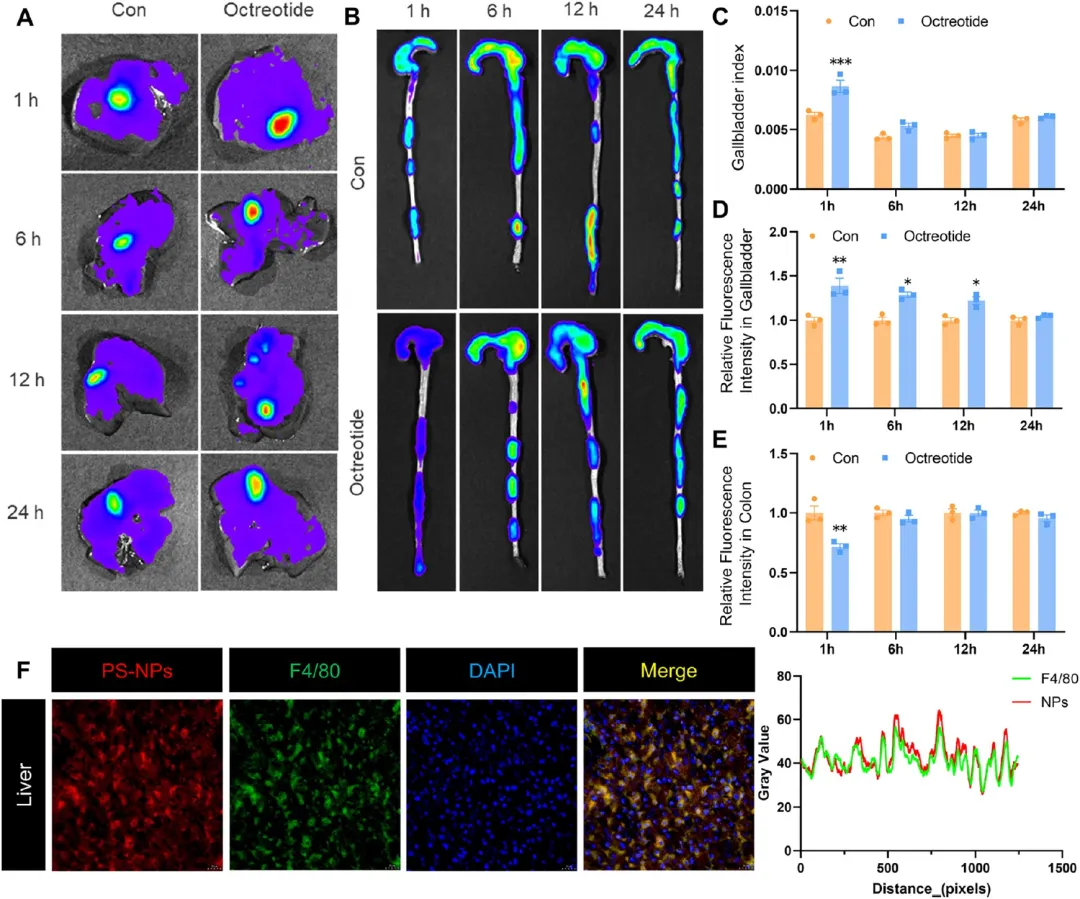
图1. PS-NPs的胃肠道吸收与肝胆排泄: (A-B) 经口灌胃100 nm PS-NPs后1、6、12、24 h胆囊和肠道的荧光图像,伴有或无奥曲肽(5 mg/kg)处理(n=3)。(C) 胆囊指数定量。(D) 胆囊相对荧光强度定量。(E) 结肠相对荧光强度定量。(F) 肝脏免疫荧光图像及线扫描,显示PS-NPs(红色)与F4/80标记巨噬细胞(绿色)共定位。奥曲肽抑制胆汁分泌后胆囊PS-NPs蓄积增加、结肠蓄积减少,证实胆汁排泄是PS-NPs进入肠道的关键途径。
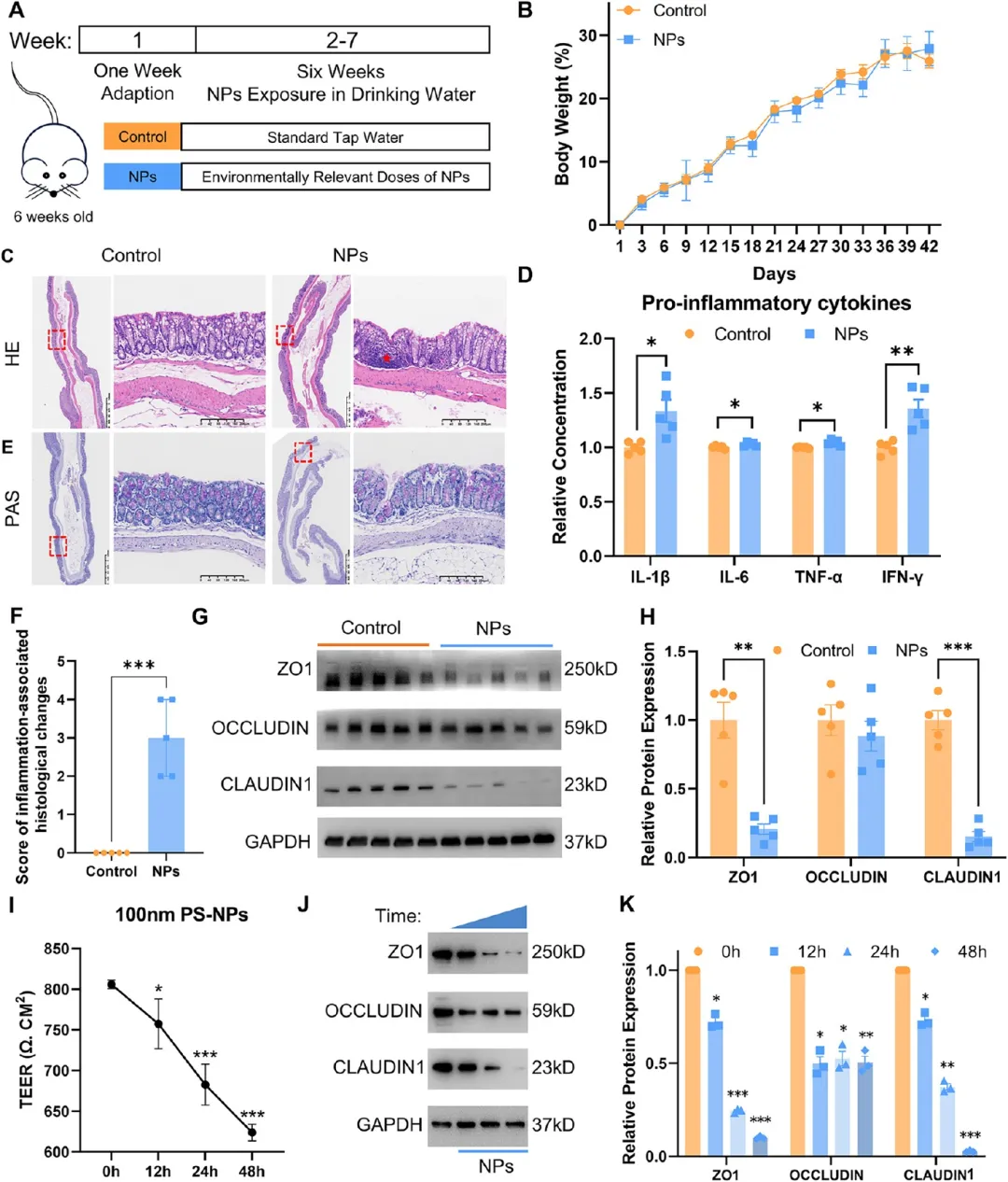
图2. 慢性PS-NPs暴露破坏肠道屏障结构并诱导炎症反应: (A) 小鼠实验流程图。(B) 体重变化(n=5)。(C) 结肠组织H&E染色代表图,标尺200 μm、1.25 mm。(D) 结肠组织TNF-α、IFN-γ、IL-6、IL-1β的ELISA定量(n=5)。(E) 结肠组织PAS染色代表图,标尺200 μm、1.25 mm。(F) 炎症相关病理变化组织学评分(n=5)。(G-H) 紧密连接蛋白ZO1、OCCLUDIN、CLAUDIN1的蛋白质印迹及光密度定量(n=5)。(I) Caco-2细胞PS-NPs处理0、12、24、48 h后TEER值。(J-K) 紧密连接蛋白蛋白质印迹(n=3)。慢性暴露后杯状细胞减少、紧密连接蛋白下调、炎症因子显著升高。
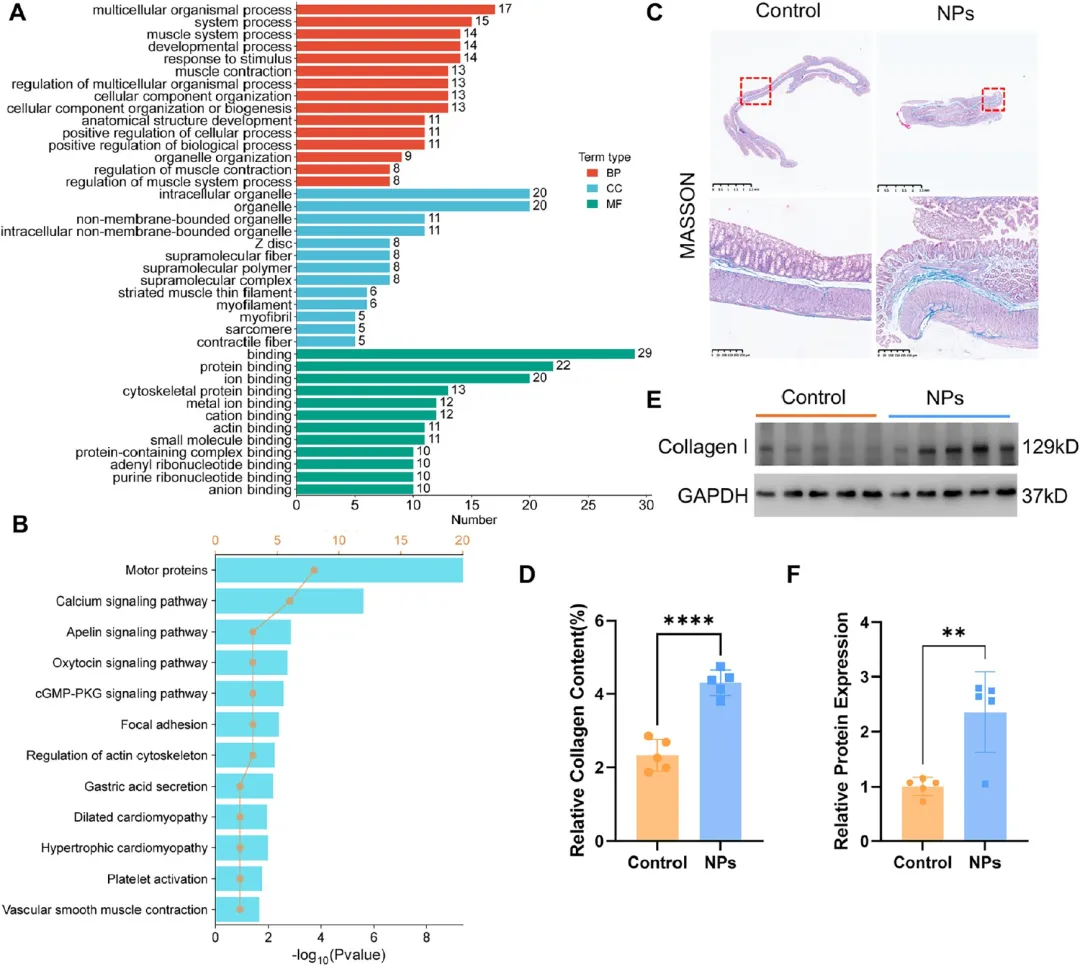
图3. 转录组学鉴定PS-NPs改变的力学转导和ECM相关通路: (A) 差异表达基因GO生物学过程富集。(B) KEGG富集分析。(C) Masson三色染色显示PS-NPs暴露结肠组织胶原沉积增加,标尺250、500 μm。(D) 胶原纤维面积定量(n=5)。(E-F) 细胞外基质蛋白Collagen I的蛋白质印迹及定量(n=5)。转录组富集于肌肉收缩调控、细胞骨架组织和黏着斑通路,胶原沉积显著增加。
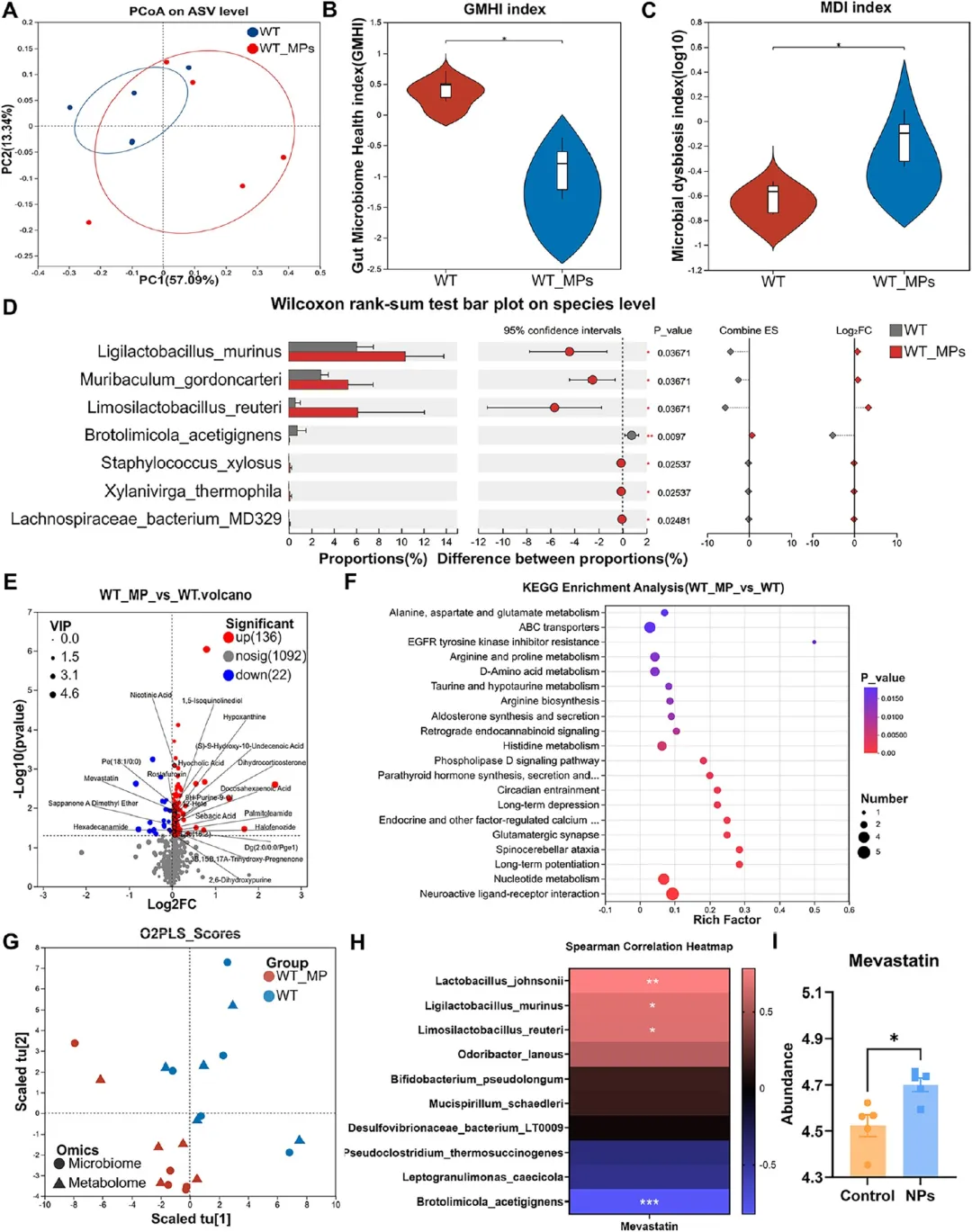
图4. 代谢组学揭示PS-NPs诱导的神经相关代谢紊乱: (A) β多样性PCoA图。(B) GMHI显著降低(n=5)。(C) MDI显著升高(n=5)。(D) Wilcoxon秩和检验鉴定的差异物种(n=5)。(E) 组间差异代谢物火山图(n=5)。(F) KEGG富集显示神经和代谢通路参与。(G) O2PLS整合分析揭示微生物-代谢物关联模块。(H) 差异微生物 taxa与美伐他汀的Spearman相关性热图。(I) 对照组和PS-NPs组美伐他汀相对丰度(n=5)。神经活性配体-受体相互作用、谷氨酸能突触等通路富集,美伐他汀显著上调。
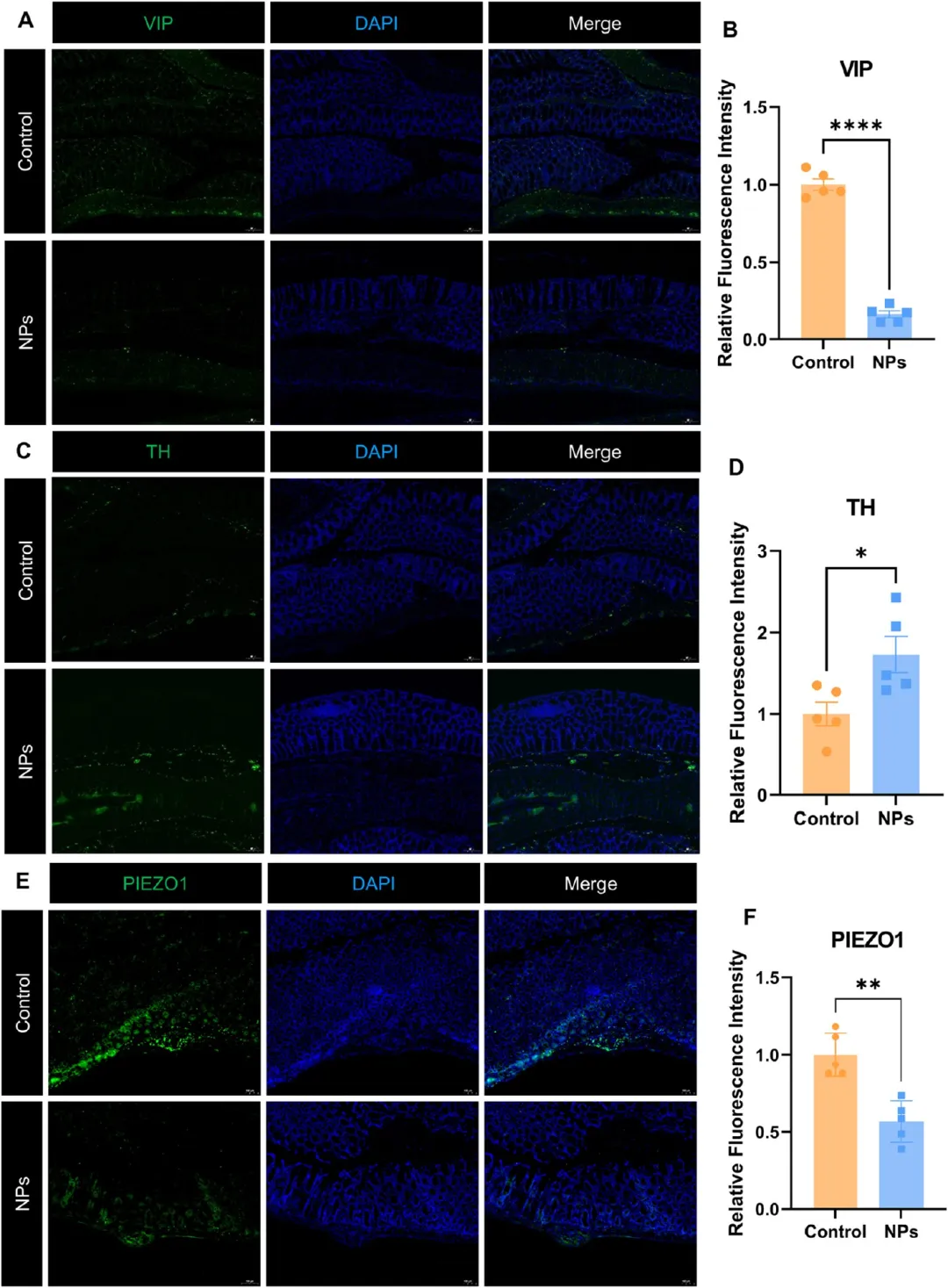
图5. PS-NPs通过破坏神经递质平衡和机械敏感信号诱导肠神经元损伤: (A-B) VIP免疫荧光图像及荧光强度定量(n=5)。(C-D) TH免疫荧光图像及荧光强度定量(n=5)。(E-F) PIEZO1免疫荧光图像及荧光强度定量(n=5),标尺50 μm。PS-NPs暴露后VIP和PIEZO1显著降低、TH显著升高,提示肠神经环路神经化学失衡。
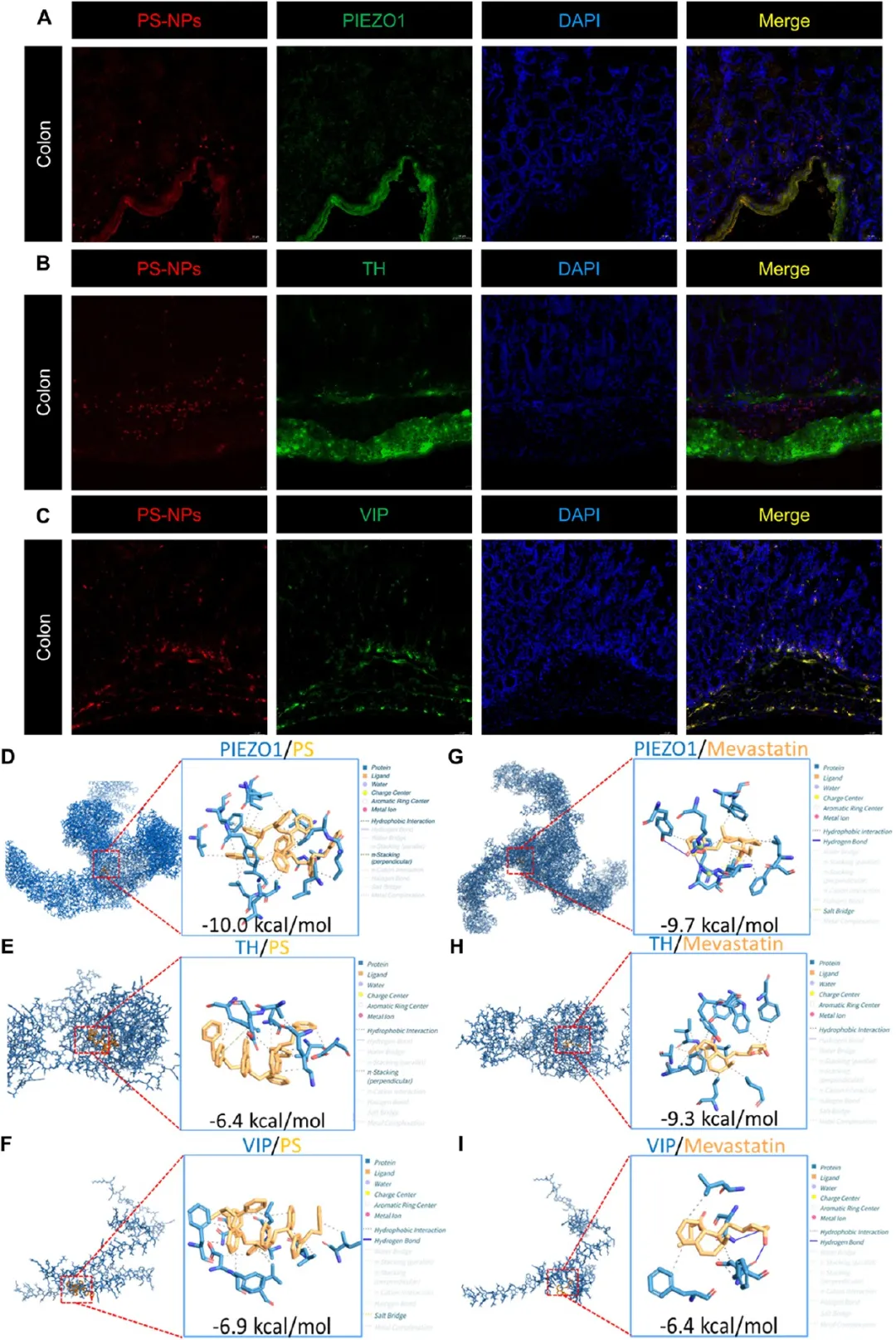
图6. PS-NPs和美伐他汀与肠神经元标志物的相互作用: (A-C) 结肠组织中PS-NPs与PIEZO1(A)、TH(B)、VIP(C)的共定位免疫荧光图像,标尺50 μm。(D-F) PS与PIEZO1(D)、TH(E)、VIP(F)的预测结合模式和结合能。(G-I) 美伐他汀与PIEZO1(G)、TH(H)、VIP(I)的预测结合模式和结合能。PS-NPs与PIEZO1和VIP呈现明显共定位及强结合亲和力,美伐他汀与PIEZO1和TH亦有强结合。
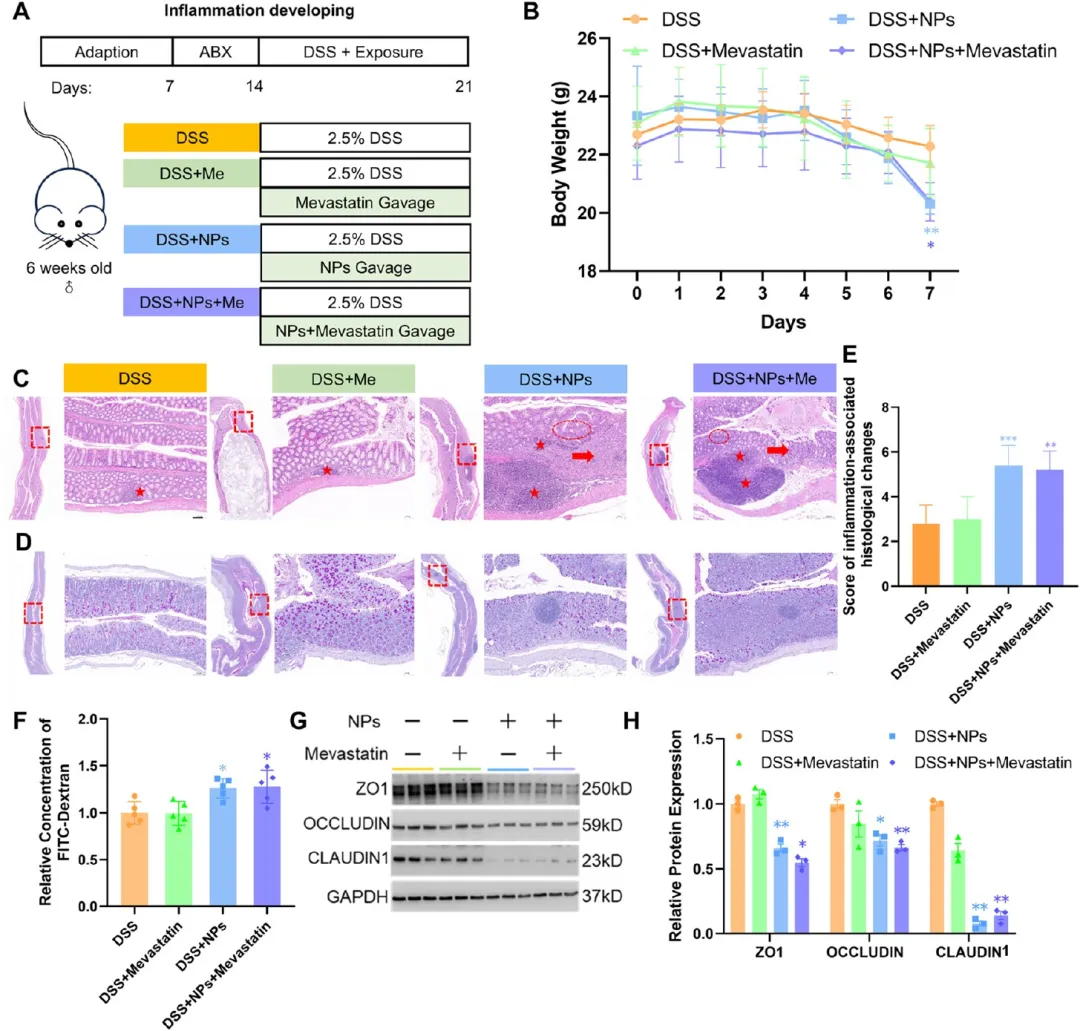
图7. PS-NPs对DSS结肠炎发展期小鼠肠道屏障恢复的影响及美伐他汀干预作用: (A) 实验设计流程图。(B) 每日体重监测(n=5)。(C-D) 结肠组织H&E和PAS染色代表图,标尺200 μm、1.25 mm。(E) 炎症相关病理变化组织学评分(n=5)。(F) FITC-葡聚糖相对浓度测定的肠道通透性(n=5)。(G-H) 紧密连接蛋白蛋白质印迹及光密度定量(n=3)。PS-NPs加重炎症损伤和体重下降,美伐他汀未能逆转PS-NPs的损害效应。
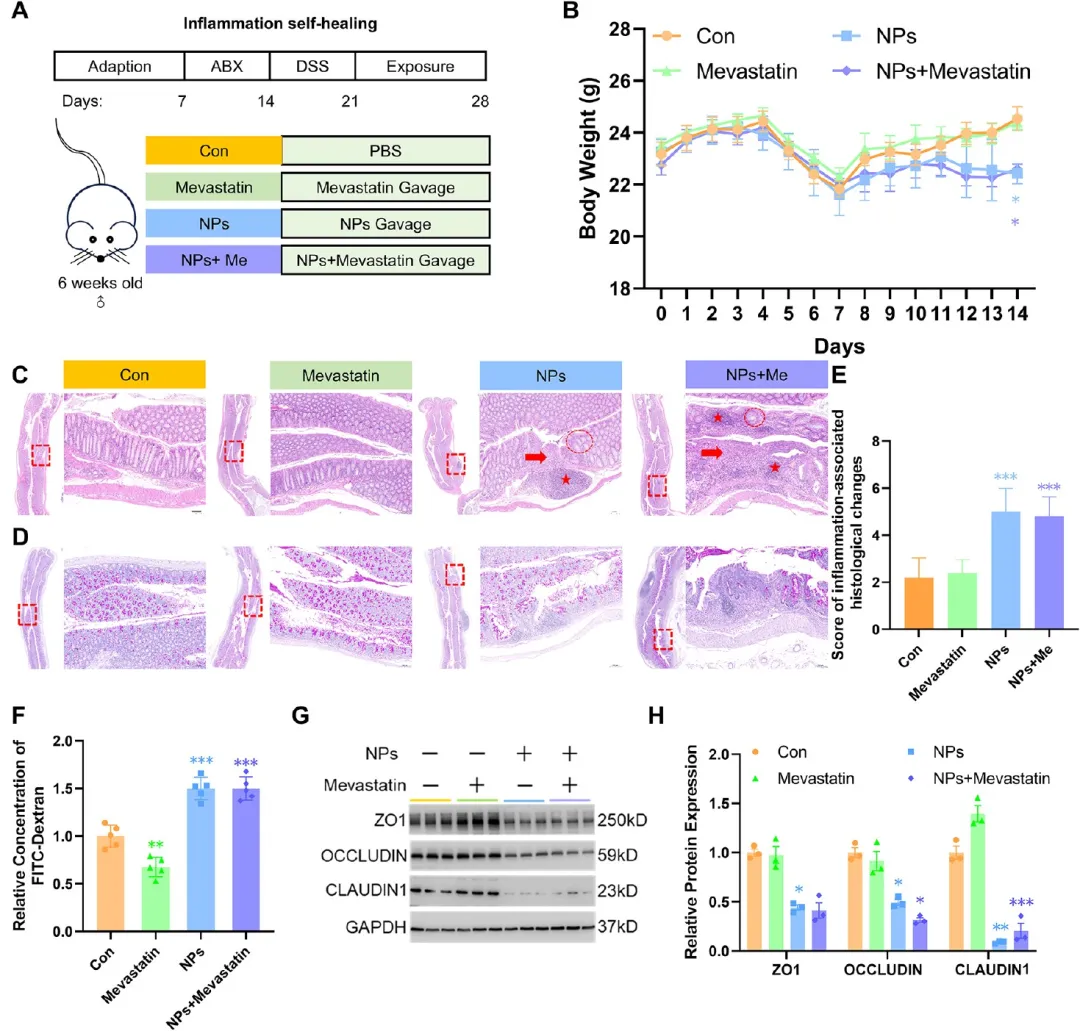
图8. PS-NPs对DSS结肠炎恢复期小鼠肠道屏障恢复的影响及美伐他汀干预作用: (A) 炎症自愈模型实验时间线。(B) 自愈期体重监测(n=5)。(C-D) 结肠组织H&E和PAS染色代表图,标尺200 μm、1.25 mm。(E) 组织学评分(n=5)。(F) FITC-葡聚糖测定的肠道通透性(n=5)。(G-H) 紧密连接蛋白蛋白质印迹及定量(n=3)。恢复期PS-NPs仍延迟屏障修复,美伐他汀单独处理显示部分保护趋势。
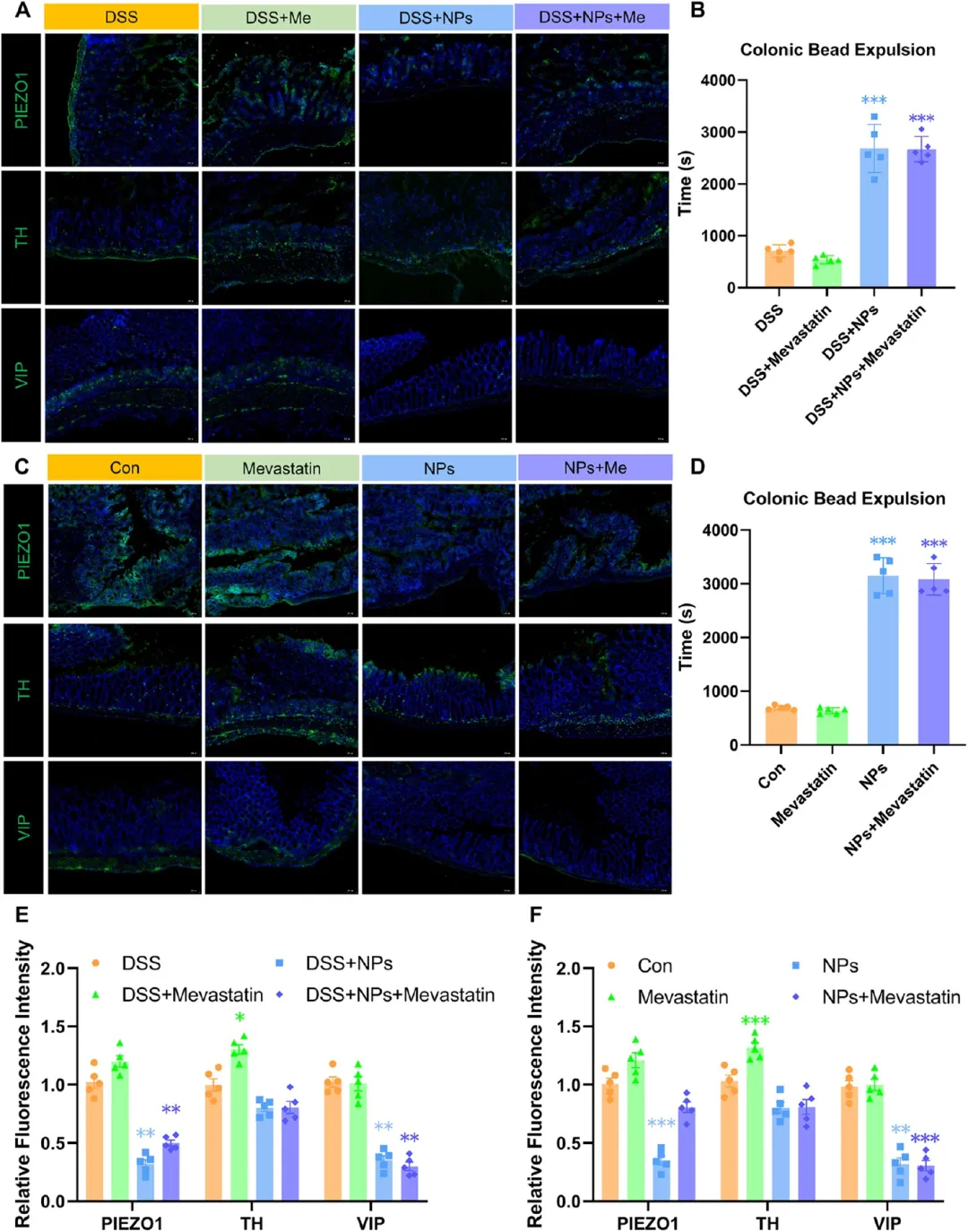
图9. PS-NPs对DSS结肠炎发展和恢复期小鼠肠道蠕动的影响及美伐他汀干预作用: (A) DSS结肠炎小鼠结肠PIEZO1、TH、VIP免疫荧光图像(绿色),标尺50 μm。(B) 急性炎症期结肠排珠时间评估肠道蠕动(n=5)。(C) 自愈期结肠PIEZO1、TH、VIP免疫荧光图像。(D) 肠道蠕动恢复评估(n=5)。(E-F) 炎症期和恢复期PIEZO1、TH、VIP相对荧光强度定量。PS-NPs暴露延长排珠时间、损害肠道蠕动恢复。
结论
本研究揭示了PS-NPs通过此前未被认识的“肝-胆-肠”肠肝循环通路驱动肠道持续暴露的机制:NPs经肠道吸收入血后被肝脏捕获,经胆汁重新排入肠腔,建立反复的肠道再暴露过程。在此过程中,PS-NPs持续破坏黏液屏障和上皮紧密连接,增加其与ENS直接接触的机会;通过下调VIP(抑制性运动神经元标志)和PIEZO1(机械敏感通道)、上调TH(交感信号),诱导ENS损伤和肠道蠕动障碍。蠕动减慢进一步延迟粪便排泄、增强NPs肠道滞留和重吸收,形成自我放大的损伤循环。菌群-代谢物分析显示美伐他汀等代谢物显著上调并与神经标志物存在强结合,提示菌群代谢物可能参与ENS调控。该研究为理解环境纳米塑料的长期肠道健康风险提供了新机制视角。
「BioMed科技」关注生物医药×化学材料交叉前沿研究进展!交流、合作,请添加杨主编微信!来源:BioMed科技声明:仅代表作者个人观点,作者水平有限,如有不科学之处,请在下方留言指正!